I gave a talk at the “Emotions in History: Boundary-Crossing Adventures” workshop, hosted by the University of California, Santa Barbara (UCSB). This symposium brought together experts from the fields of psychology and history to explore the interplay between emotional theories across these disciplines.
The talk also included a wonderful roundtable discussion.
For those interested, you can watch the recordings of this event, accessible here. My deepest thanks to UCSB and the organizers, Hongbo Yu and Ya Zuo.
As the Co-founder and Technical Director of the DRH, I’m excited about the collaborations and applications of recent advancements and the integration of novel methodologies in the study of religious history.
I spoke about “Mapping Psychological Terrae Incognita: Explorations Beyond WEIRD Psychology”, which primarily focused on these papers:
Muthukrishna, M., Bell, A. V., Henrich, J., Curtin, C., Gedranovich, A., McInerney, J. & Thue, B. (2020). Beyond Western, Educated, Industrial, Rich, and Democratic (WEIRD) Psychology: Measuring and Mapping Scales of Cultural and Psychological Distance. Psychological Science, 31(6), 678-701. [Download] [Supplementary] [Code] [Summary Post] [Publisher] [Twitter]
White, C. J. M., Muthukrishna, M. (equal senior) & Norenzayan, A. (2021). Worldwide evidence of cultural similarity among co-religionists within and across countries using the World Values Survey. Proceedings of the National Academy of Sciences, 118 (37) e2109650118. [Download] [Supplementary] [Publisher] [Twitter]
Muthukrishna, M., Henrich, J. & Slingerland, E. (2021). Psychology as a Historical Science. Annual Review ofPsychology, 72, 717-49. [Download] [Publisher] [Summary Post] [Twitter]
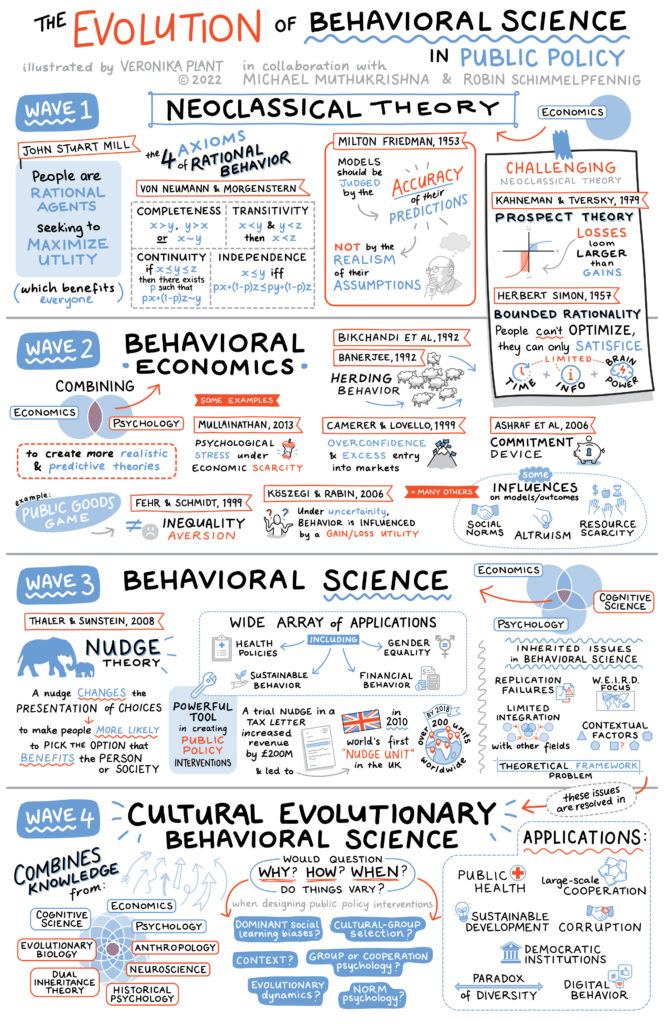
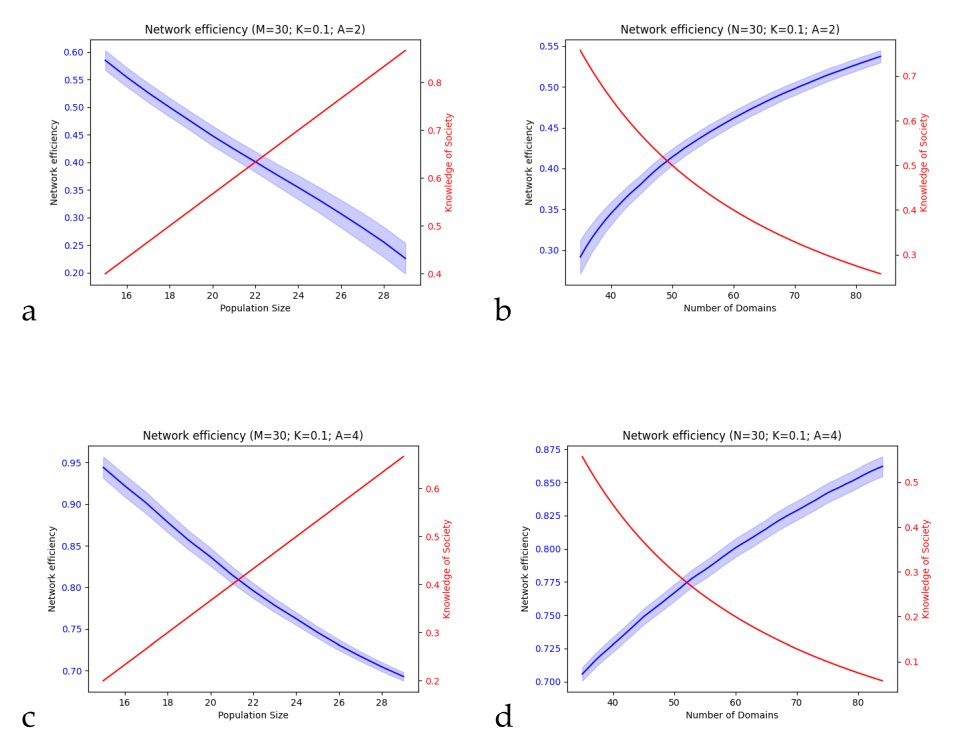
My talk begins with an overview of the problem plaguing behavioral economics – the lack of a theoretical foundation that can guide policy interventions. I introduce cultural evolution as a possible solution to bridge the theoretical gap. By using cultural evolution as a theory of human behavior, improvements can be made in policy efficiency. For example, studying how social norms change and evolve over time will provide a foundation for implementing effective policy interventions in multicultural societies. Some of this history is captured in this figure from the paper:
Considering the historical path dependence of norms provides crucial in understanding why certain populations hold certain beliefs, like vaccine hesitancy and a distrust in healthcare systems. Identifying how people acquire cultural norms, and narrowing down the ultimate causes for behavior (through cultural distance tools like world.culturalytics.com) could provide insights into designing interventions that work.
Understanding cultural evolution and behavioral science can help reanalyze the literature on public policy, providing insights into why some approaches are successful while others are not. I explain how studying universal cognitive capabilities will provide a deeper understanding of norm change, and thus, improve policy design.
Robin Schimmelpfenning and I discussed our co-authored paper on the paradox of diversity in the collective brain. We discuss how the rate of innovation depends on sociality, information fidelity, and cultural trait diversity. While cultural trait diversity offers the largest potential for empowering innovation, it also brings with it potential coordination and cooperation challenges. Diversity, in other words, is both a source of innovation and divisive; a double-edge sword. We then propose using cultural evolvability as a framework for resolving this paradox.
Our presentation was followed by a discussion session and subsequently a panel discussion chaired by Dominic Abrams.
Here’s the take home: diversity empowers innovation through recombination but also by definition divides us. We call this the paradox of diversity. A principled way to resolve this tension is by considering cultural evolvability.
We discuss implications for entrepreneurship, polarization & a nuanced take on diversity. This framework can also guide researchers and practitioners in how to reap the benefits of diversity by reducing costs.
Let’s start with innovation: A folk understanding of innovation is that it’s driven by a talented few – the giants upon whose shoulders we stand. But that view is inconsistent with theoretical and empirical work in cultural evolution.
Instead, innovations emerge as ideas flow through our social networks, requiring a specific innovator no more than your thoughts require a specific neuron. See: Innovation in the Collective Brain
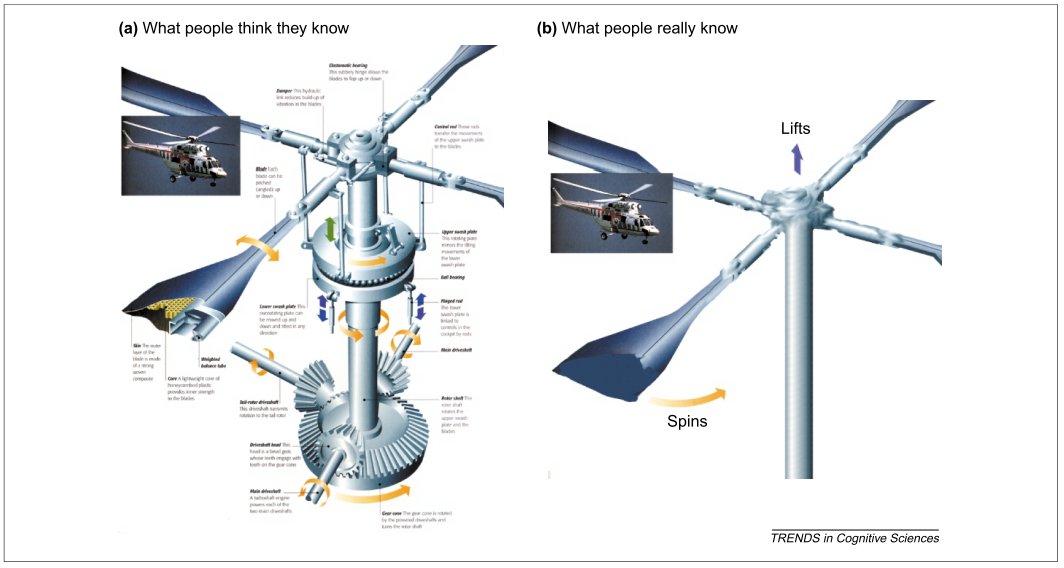
People are often unaware of how little they actually deeply understand about the world – what’s referred to as the “knowledge illusion” or “illusion of explanatory depth”. The Knowledge Illusion is a good pop book on the topic.
The world is not only complicated, but more complicated than our psychology allows us to believe. Innovations emerge through incremental improvements through partial causal models and large leaps through serendipity & recombination.
Three key levers that affect innovation are sociality (size & interconnectedness of a population), transmission fidelity (how well you can transmit information between people), & cultural trait diversity. Diversity has the most potential benefit and the largest challenges.
Let me say a bit about each. 1. Sociality: in general +ve relationship, because large pops had to solve the coordination problem to become large. Interconnectivity has an optimal point. Small work groups can be easily overconnected; large pops should be more connected.
2. Transmission fidelity: under selection as cultural complexity increases. Hunter-gatherers not much explicit instruction.Industrial revolution eventually led to schools to download a minimum common cultural package – reading, writing, arithmetic, algorithms for thinking.
Today we have the printing press, radio, TV, Internet, social media, and Zoom. But there’s still too much to learn. Unless you get a PhD, 21C students don’t learn mathematics developed after 1900; scientific training is longer, & major contributions are made at an older age.
We spend longer learning, delaying kids. Theres limit to pop size & transmission; how long you can delay being productive. Another path is to divide up info & labor-specialize. Get smart at 1 thing & stupid at everything else: cultural trait diversity
Different kinds of diversity & different ways to measure it. Our focus is on cultural trait diversity-beliefs, behaviors, assumptions, values, technologies, & other transmissible traits. e.g. languages, processing techniques, technical skills, family structure & occupation.
In the public discourse, diversity often refers to ancestry or physical characteristics-skin color, ethnic origin, religion, sex, gender, sexual orientation, or ability. These may correlate with cultural trait diversity, but correlation may weaken over generations.
For example, Americans with different ancestries may possess similar WEIRD psychology (part of why I have an issue with the WEIRD=White take, though that’s a separate issue). I’ll use diversity from herein, but that’s what we mean.
Diversity can be distributed in different ways: Diversity between pops culturally evolves as pops adapt to local differences, influencing future generations through historical path dependence created by past conditions or founder populations. See also: Psychology as a Historical Science
Diversity within populations evolves as information and labor are divided as discussed above.
Within-population diversity includes disciplinary differences, such as the sciences and humanities, industry specialisations, guilds, and firms. Diversity can be structured as ‘cultural clusters’ by ethnicity, class, wealth, occupation, politics, religion, or incidental geographic layout. Cultural clusters may intersect, such as in ethnic occupation specialization-lots of examples. See also: Beyond WEIRD Psychology: Measuring and Mapping Scales of Cultural and Psychological Distance
Diversity may also exist within certain individuals—multicultural individuals, ‘third culture kids’ (like Barack Obama), interdisciplinary researchers, and so on.
Diversity is therefore both the product of cultural evolution and fuel for the engine of further innovation
But without common understanding & common goals, the flow of ideas in social networks is stymied, preventing recombination, and reducing innovation. Consider communication without a common language or collaborations between scientists & humanities, or different scientists.
We formalize this paradox of diversity trade off in the paper and will be exploring solutions in future work. Check out the paper for details. We argue cultural evolvability is the right way to think about this.
Evolvability refers to not how well adapted a population is to current circumstances, but its ability to evolve to future circumstances. Variation or diversity, and the forces that create and stabilise that diversity are key factors that create evolvability.
Cultural evolvability is a balance between diversity and selection, exploring and exploiting, sampling and specialising, convergent and divergent thinking, stability and change, efficiency and flexibility.
Lots of related work on diversity and selection, explore-exploit or sampling-specialising trade-off in development, search for global solutions & avoidance of saddle points within machine learning.
As an aside, ML insights: in a sufficiently high dimensional space there are effectively no local optima, only saddle points with some dimension that allows escape. Biological & cultural systems have large dimensionality, there are no true evolutionary stable equilibria.
Next, we review the diversity literature through the lens of cultural evolvability. First we lay out 9 challenges to interpreting the literature. It’s a literature that would be benefit from theory. See also: A Problem in Theory
Diversity overlaps with challenging aspects of psychology, norms & institutions: racism, prejudice, xenophobia, sexism, discrimination, power differences, social and economic inequalities.
Love to write about at some point, here we focus on coordination challenges, which influence and are influenced by these problematic features of the world. Our goal is to review the overall patterns in the literature and make sense of these in light of cultural evolvability.
Here’s some of the mixed lit: Within countries, diversity is often approximated by birthplace diversity, professional diversity, ethnic diversity or linguistic diversity. Research looking at the relationship between diversity and economic growth suggests: positive effect of birthplace diversity, but -ve effect of ethnic and linguistic diversity. Research asking questions about immigrants in general often ignore the heterogeneity – cultural distance and education (effectively the cultural traits in your head) matter. Even more mixed in firms. Overall, educational diversity and deep level diversity seem to be positive for innovation within a firm. Mixed effects within teams – we think the paradox of diversity can disentangle.
We derive some insights:
1. Cultural evolvability means tolerance for diversity. Currently less adaptive traits may be more adaptive when the environment changes. Different environments lead to different evolvability strategies. In materially insecure societies, not following a successful “Tiger Mom” strategy—working hard to secure scarce educational opportunities and subsequent employment opportunities—has a much larger cost. But this leads to incremental over radical innovation.
2. Cultural evolvability means under-optimization & inequality. Cultural evolvability necessarily means inequality in outcomes, because not all will have the optimal strategy for the current environment.
Firms face a tradeoff: strategies that favour efficiency & strategies that favour flexibility. Consistent, strong cultures perform well in stable markets, but poorly during times of change. Under-optimizing and allowing for flexibility increases a firm’s evolvability.
Not all firms can bear the cost of under-optimising in the short term-high risk, high value approaches better suited to larger firms or larger countries. Read about Satya Nadella and Microsoft:
Compromize strategies: skunkworks, ecosystems of different firms trying different strategies (e.g. Silicon Valley), or countries composed of different states or regions trying different approaches (e.g. what Justice Brandeis described as “laboratories of democracy”).
In shared multi-agent reinforcement learning, diversity increases performance through exploration and individualized behaviors. Evolvability means many approaches will be suboptimal or even fail, but the successful approaches can be spread and benefit the group as a whole.
Indeed one of the benefits of access to multiple cultures in pluralistic, multicultural societies is the ability to create new approaches by learning, borrowing, copying from each other and other cultures. We should do more of that.
3. Cultural evolvability helps explain levels of entrepreneurship. Cultural evolvability requires doing something different. Most new businesses fail & the willingness to take a risk depends on personal and population-level costs and benefits.
A. personal cost of deviation: many deviations will result in lower payoffs than following the majority trait. If it were obvious how to do better, most of the population would already use the better strategy. Tolerating diversity in traits, thus, means tolerating failure.
Reducing cost of failure increases entrepreneurship: bankruptcy laws, social safety nets, rich parents – a child with parents in the top 1% income distribution is 10 times more likely to be an inventor than a child born below the median, controlling for measures of ability.
B. population-level benefit of deviation. In a large economy with a large customer base comes large rewards for large innovations – the few winners can win bigger. Amazon can make more money in the United States than in Australia. Here’s a great video of Bezos describing his vision back in 1997:
C. Who pays the cost and who benefits from the innovation at a population-level, a function of the scale of cooperation. Even if at an individual-level the benefits of entrepreneurship don’t outweigh costs, they may do so at a population-level.
Silicon Valley offers an example. For every Apple & Amazon, there are 1000s of start-ups that have failed – most start-ups fail & the overwhelming majority never receive funding (114) – ‘unicorns’ are called unicorns for a reason. But the few successes pay for the failures.
4. Cultural evolvability prevents polarization & cultural speciation. Harshly punishing minor deviations increases extremism. If you’re harshly punished anyway, may as well take the extreme position. If there weretolerance for diverse view points, I’d moderate my position.
Model and results also inform debates on freedom of speech, predicting large sanctions for small deviations may encourage a divided society. By corrolary tolerating multicultural diversity of opinions and cultural traits may prevent polarization.
Cultural clustering complicates everything, but I’ll let you read about that in the paper. It gets into colonialism, resource competition, and intergroup violence. 50/ To conclude, diversity has been central to the success of all life. Until around 1.2 billion years ago the source of that diversity was mutation – genetic innovation through serendipity and incremental improvement alone. Single cells reproducing by simple replication.
Sex unlocked the recombinatorial power of diversity, increasing evolvability and the speed of evolution. So too with culture, but there are many barriers to cultural traits meeting and recombining.
We live in an increasingly connected & multicultural world. Migration is a constant feature of the human story, but since the Age of Mass Migration, more people from more culturally distant societies live side by side. Their countries of origin must coordinate as never before.
So much human potential is lost through unequal access to information and adaptive cultural traits. The goal of any society or org should be to reap the benefits of diversity and minimize the costs, thereby maximizing human potential. We discuss several strategies.
Humans are a deeply cooperative species. Our greatest achievements and our worst atrocities are both cooperative acts. In a more diverse world, the challenge is greater, so too are the potential gains.
Long post, but an important topic that helps to resolve controversies such as IQ differences.
Quick summary: We reconcile behavioral genetics and cultural evolution under a dual inheritance framework. A cultural evolutionary behavioral genetic approach cuts through the nature–nurture debate and helps resolve controversies such as IQ.
Business is booming in behavioral genetics. We’re in the midst of a GWAS gold rush. Powerful computers and sequenced DNA of millions has led to an industrious search for SNPs that correlate with a variety of traits. Some even claim the curse of reverse causality has been lifted.
There’s also been a parallel revolution in cultural psychology and cultural evolution. Genes, culture, and the environment have often co-evolved, shaping our species. But the revolutions in behavioral genetics and cultural evolution have largely been independent. But, given the extensiveness of the cultural and culturally-shaped environment, cultural evolution offers an important but typically missing complement to otherwise insightful methodological and empirical analyses within behavioral genetics. Genes and culture are intertwined. For example, our jaws too weak and guts too short for a world w/o controlled fire & cooked food. It’s obvious that lower environmental variation will lead to higher heritability scores. Less obvious is how culture can mask or unmask genetic variation. Or how diffusion and innovation can increase or decrease heritability. Or how to define a single society for the purposes of measuring heritability, without being able to identify cultural cleavages that can lead to Scarr-Rowe type effects: en.wikipedia.org/wiki/Scarr-Row…
Reconciling behavioral genetics & cultural evolution offers insights for differences in heritability between and within populations, differences in heritability across development, and the rise in IQ (Flynn Effect). We’d like a discussion that nuances common interpretations of the nature and nurture of behavior.
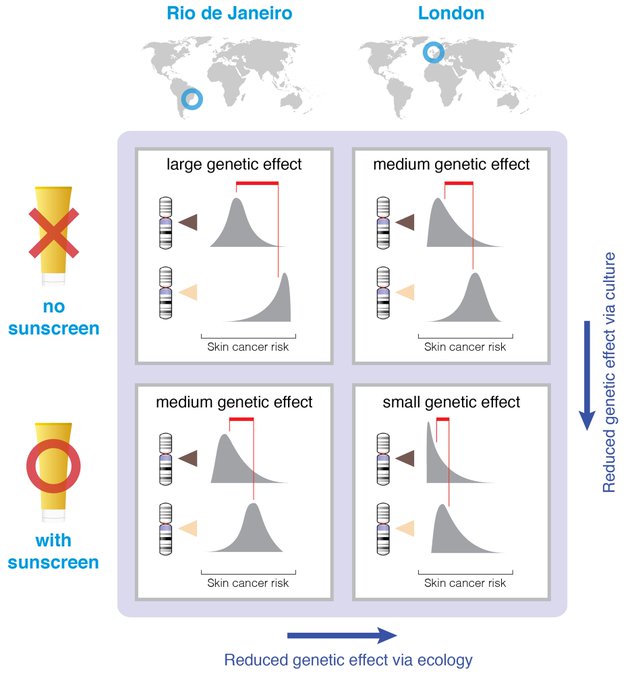
First, let’s quickly get a common misinterpretation of heritability out of the way: Heritability is not an index of the genetic basis of a trait nor a measure of the relative contribution of nature compared to nurture. It’s the proportion of phenotypic variance for some trait that is explained by genetic variance. So obviously variability in genes, in environment, and in traits will all matter. A quick illustration: skin pigmentation and UV levels. Genes affect level of skin pigmentation and propensity for tanning instead of burning; ancestral adaptations to UV radiation at different latitudes.
Migration means melanin is mismatched to latitude: Aussies with European ancestry are more susceptible to skin cancer; Europeans with African and South Asian ancestry have higher rates of vitamin D deficiency.
A Gene×Environment approach won’t predict how heritability estimates change over time as non-genetic adaptations compensate for genetic mismatches: fairer Australians wear sunscreen, a hat, & covered clothing; darker Europeans consume vitamin D supplements & vitamind D-rich or fortified foods. Here it’s easier to see that heritability is a function not only of genes, traits, and ecology, but also of an evolving cultural environment. The environment is not an inert backdrop against which genes should be evaluated. It evolves in relation to both genes and ecology.
Four lessons before we continue. The first is obvious to behavioral geneticists, the second sometimes noted, the third and fourth are typically missing.
Lesson 1
There is no overarching, one-quantity heritability of a trait to be discovered. There is no fixed answer to the question, “What is the heritability of skin cancer?”
Lesson 2
Heritability will depend not only on ecology, but also on culture & specifically on diffusion and innovation—both of which can rapidly change and therefore rapidly change heritability estimates.
Lesson 3
Diffusion and innovation are broadly directional. Cultural diffusion of sunscreen, clothing, shade & sunglasses, and cultural innovation toward more effective screening and treatment of melanomas all work to reduce heritability estimates due to a masking effect.
Were any of these an example of culture unmasking genetic effects, such as tanning salons that induce differential risk according to skin pigmentation level, we would have predicted an increase in heritability.
Lesson 4
We might expect a stronger cultural response where ecological and cultural selection pressures are stronger—skin cancer mitigation in Australia but Vitamin D supplementation in northern Europe. Not been tested to our knowledge, but the predictions are clear.
“Which SNPs are associated with skin cancer?” is similarly culturally dependent. Societies where sunscreen use is common, we expect SNPs associated with skin pigmentation to be less predictive of skin cancer compared to societies where this is not the case.
Similarly, we would expect SNPs associated with antioxidant metabolism to be less predictive of skin cancer in societies whose foods are rich in antioxidants—such as in traditional Mediterranean cuisine.
Section 2.2 is on how cultural evolution shapes heritability through diffusion and innovation. We live on the peaks climbed by cultural evolution – human environments have already been shaped by cumulative cultural evolution—functionally overlapping with genetic evolution.
Diffusion and invention can mask or unmask genes. Examples using language, fertility, and schooling.
Language example
If Cantonese or Yoruba (both tonal) spread, heritability of language ability would increase proportional to variation in “tone” genes.
If Norwegian or Russian (both non-tonal) spread in the same population, heritability of language ability would decrease.
Fertility example
Contraception and social values in 20th century unmasked the effect of genes associated with reproductive behaviors and preferences (heritability rose in US). But a one-child policy or rigid childbearing norms masks the genetic effect.
School example
School is a powerful mechanism for cultural diffusion. Heritability of literacy in:
Australia: Kindergarten: 0.84 Grade 1: 0.80
Scandinavia: Kindergarten: 0.33 Grade: 0.79
Why?
Cultural diffusion of literacy.
Australian children begin receiving compulsory literacy instruction in kindergarten, while in Scandinavia the kindergarten curriculum emphasizes social, emotional, and aesthetic development—literacy instruction only begins in Grade 1.
Assessing the genetic basis of literacy without accounting for particulars of curricula on cultural diffusion is a selection bias of unknown magnitude. Note that literacy in the home environment is already shaped by cultural evolution; there is no ‘baseline’ heritability. Heritability is a composite measure that captures both genes and culture. Saying literacy heritability in Scandinavia jumps up to 0.79 in Grade 1 reveals as much if not more about the disseminative power of modern schooling than it does about the genetic basis of literacy. Similar dynamics with innovation. Read about it in the paper.
However, one neglected factor is “cultural clustering”, where even highly useful forms of cultural knowledge may not easily permeate social barriers. Not necessarily ethnic boundaries, also class, wealth, occupation, political alignment, religion, or incidental geographic layout. Greater differential clustering can lead to a cultural Simpson’s paradox (we’ll get to that shortly, but see Section 3.4).
Cultural FST (CFST) is useful for identifying these clusters. You can read more about that paper here: https://www.michael.muthukrishna.com/beyond-weird-psychology-measuring-and-mapping-scales-of-cultural-and-psychological-distance/
Hopefully, you can see the importance of a cultural evolutionary behavioral genetics. We hope this target article will spark a vibrant discussion. But let’s move onto the problems that obscure the effect of culture:
(1) the WEIRD Sampling Problem
(2) the Hidden Cluster Problem
(3) the Causal Locus Problem
And then describe the:
(4) Cultural Simpson’s Paradox that emerges at their junction.
The WEIRD Sampling Problem
The WEIRD people problem? Pretty bad in genetics too. Twin studies: 94% Western: 60% US, UK, Aus; 25% Nordic 6% Non-Western: 4% China, Japan, South Korea, Taiwan
Remainder of the world, i.e. vast majority of humans are the remaining 2%
Same story in GWAS: 88% European ancestry. 72% from just 3 countries: US, UK, & Iceland 20% from Japan, China, and South Korea
From a cultural evolutionary perspective, given (a) cultural environment, (b) coevolution b/w culture & genes, & (c) cultural differences between populations, not surprising that: 1.Polygenic scores don’t translate well across ancestry groups (European scores, 42% in Africa) 2. Polygenic scores are highly sensitive to inadequately controlled population stratification. And so cultural variation and the hidden cluster problem is pernicious.
Hidden Cluster Problem
Cultural clusters (or segregated diversity) typically created by barriers impeding cultural transmission, such as topography, cultural conflict, language, social stratification by class, wealth, etc. Immigrant countries more clustered (Canada > Japan).
Countries whose borders are drawn arbitrarily with respect to the geographic arrangement of cultural groups, for example by colonial administration (many countries in Africa), are also likely to have high clustering. You can use CFST to find them: https://journals.sagepub.com/doi/abs/10.1177/0956797620916782
Note that cultural clustering is not the same as genetic clustering as we explain at length in Section 3.2.2. Indeed, reconciliation between cultural evolution and behavioral genetics requires an update in the way we think about culture.
Causal Locus Problem
Hidden cluster problem describes complexity that exists w/in social groupings. Culture is not an unstructured exogenous variable. Culture is constructive system that accumulates functional adaptations in a directed manner over time. Two key lessons here.
Lesson 1: Genes that make vs genes that break. The more complex a system, the more ways it can fail. Take the history of lighting.
Wood fire can be extinguished in 2 ways Flourescent bulbs have 7 ways to fail LEDs have 30
Faulty O-ring can explode a space shuttle and so on.
There is a fundamental asymmetry: easier to find ways to break the system than ways to explain or improve it. So too for gene function. All your cells have the same bootstrapped code, but they interact with each other, what they create, and their surroundings to create you.
There are many ways these interactions can go wrong. It is easier to identify deleterious genetic mutations than beneficial mutations. The space of failure is larger than the space of success, making genes that break more detectable than genes that make. For example, a single mutation can cause Mendelian disorders such as cystic fibrosis and Huntington’s disease, but no single mutation creates genius. Over 1000 genes have been linked to intelligence.
Each gene only explains a miniscule fraction of variation in intelligence, and the causal mechanisms are unlikely to be straightforward. In contrast to these genes that make, the causal mechanisms behind single gene mutations that cause intellectual disability—e.g. BCL11A, PHF8, ZDHHC9—are relatively well understood.
Increasing nutrition, improving schooling, and removing parasites have positive effects on IQ, but in a society where parasite infection is kept under control, we would not notice that parasite status correlates with intelligence. And by corollary, genes that provide protection against malnutrition, parasites, or pollution would only be positively associated with intelligence in environments where these insults occur. In environments where these insults have been removed, the same genes would not be associated with intelligence, and can even be deleterious, as with sickle cell trait. Not helpful if there’s no malaria.
Genes are functionally masked by cumulative cultural evolution, and we expect that this masking is extensive and systematic. A quick evolutionary and historical example: Vitamin C, the GLO gene, and dead sailors.
Vitamin C is an essential nutrient and its acquisition is thereby an essential biological function. Endogenous synthesis of vitamin C requires a gene called GLO, and GLO is present across most of the animal kingdom. But because vitamin C synthesis is metabolically costly, the gene is inactive in some species that have access to sufficient quantities of the nutrient in their diets. e.g. taxa such as teleost fishes, guinea pigs, many bats, some passerine birds, monkeys and apes.
Anthropoid primates occupy a frugivorous niche, and fruits often contain sufficient vitamin C. Here gene function is offloaded onto environmental resources. In turn, this offloading has behavioral implications. If a species becomes dependent on its environment for vitamin C, both its behavioral range and evolutionary trajectory become constrained by the availability of the nutrient. Humans are a nice example of this.
As our species migrated across the planet, we found ourselves in environments where vitamin C was in short supply. A deficiency of vitamin C causes scurvy—the bane of seafarers until the trial-and-error discovery that certain food items like sauerkraut and citrus could prevent ships from being packed with tired, bleeding, toothless, and eventually dead sailors.
Masking does not necessarily need to be in the direction from culture to genes: genetic assimilation is same process working in the opposite direction, where a trait that is regularly acquired through learning gradually transfers its locus to the genome (i.e. Baldwin effect).
Cultural Simpson’s Paradox
Which leads us to the Cultural Simpson’s Paradox. Causal Locus Problem can confound the measurement of genetic effects due to Hidden Cluster Problem obscured by WEIRD Sampling problem creating a Simpson’s paradox.
Let’s return to the UV example. The melanin-UV mismatch can be masked by the cultural diffusion of sunscreen, especially in regions with more exposure to sunlight. In other parts of the world, the issue is under-exposure to the sun causing vitamin D deficiency. Low vitamin D leads to lack of bone integrity, muscle strength, autoimmune disease, cardiovascular disease, cancer etc.
In US and France, more north you go, the the lower vitamin D levels. Makes sense, right?
But when we compare across Europe, we see the opposite pattern where people in northern countries have higher vitamin D than people in southern countries. What’s going on?
High consumption of fatty fish and cod liver oil in Northern Europe, as well as greater sun-seeking behavior in these countries compared to Mediterranean Europe. These are potent cultural adaptations.
Participants fed the traditional Norwegian fish dish mølje three times over a span of two days had 54 times the recommended daily dosage of vitamin D. The relationship between latitude and Vitamin D goes one way within a country, and the other way between the countries.
If we had been Martian anthropologists who did not know that the populated landmass known as “Europe” can in fact be broken down into sub-units called “countries”, these examples would be standard examples of a Simpson’s paradox.
In these cases, the paradox occurs when we do not know how to partition the higher-order population (Europe) into lower-order units. Fortunately, we can partition continents into countries, but in other cases, the relevant units is not as easily identifiable. Let’s move on.
We now have enough to make sense of puzzles in behavioral genetics such as (1) differences in heritability across socioeconomic levels, (2) differences in heritability across development, and (3) the Flynn effect.
SES: Heritability of IQ is higher among affluent, high socioeconomic status (SES) households than among poorer, low-SES households in some societies, but mixed in others. Why?
One explanation is ‘reciprocal causation’: genes well suited to a task can better nurture their skills in a wealthier environment than in a poorer environment and this is amplified over time. Maybe, but then why don’t we see the effect in Europe and Australia?
Here’s what we think is going on: in the US, the differences between school and home environments among high-SES households is smaller than among low-SES households. US is a land of variance. Factors such as school lotteries can dramatically affect the cultural input.
In contrast, the cultural environment is less unequal in western Europe and Australia, where, for example, high quality schools are available across SES. Where these two explanations make different predictions is for poorer countries.
Reciprocal causation would predict low heritability in poorer countries. We would predict high heritability where there is equal access to similarly poor schools and household conditions, but low heritability if inequality is high.
Incidentally we predict the opposite between human and animal environmental effects due to social transmission. It’s interesting, but not central. Check out Section 4.1.2. Let’s move onto heritability across development.
Heritability changes over the lifespan. Heritability of political orientation is similar for American identical and fraternal twins from middle childhood up to early adulthood. Right around the age at which American children leave home, this pattern is broken.
Drops for fraternal but not identical. We argue this is due to vertical vs oblique transmission and would predict a different drop off for say Italian or Croatian who leave home past 30.
Flynn effect describes the rise in IQ test scores over time. Largest in countries that have recently started modernizing, and smallest in countries that had attained modernization. No consensus to explain it, but given speed genes obviously unlikely.
We argue its caused by a rapid worldwide increase of cultural practices, technologies. Intelligence is about hardware—genes, parasites, pathogens, pollution, and nutrition affecting health and brain development, but also software—our increasingly complex cultural package.
By this account, not only is the idea of a culture-free IQ test implausible, but so too is the idea of culture-free IQ. Lots to say here. Go read Section 4.3.
Home stretch: Cultural Evolutionary Behavioral Genetics. The thrust of our theoretical case is that human psychology and behavior have a large cultural component that has been changing over history.
Most recently our psychology has been shaped by the advent of writing, numeracy, different types of agriculture, the Industrial Revolution, the Internet, and smart phones.
As new adaptive traits emerge, initially those who possess these traits will have an advantage, as in the case of access to new food sources, better healthcare, more efficient technologies, or easier methods of learning.
But eventually the traits will reach fixation in the population through the processes of cultural diffusion, at least until they are unseated by subsequent innovations. We predict that these cultural dynamics are reflected in heritability estimates.
As any geneticist knows, genetic heritability is a function of the variability in the environment, variability in genes, and variability in the phenotype. There is little to predict if the phenotype is homogenous, as in the number of fingers or kidneys.
There is little to predict with if the environment or genes are homogenous. But what is factored into the environment includes not only the physical ecology, but also the cultural environment.
While variance in genes and ecology may be relatively stable, the variance in the cultural environment is continually changing through the processes of cultural evolution. A genetic account of human psychology and behavior must also account for culture and cultural evolution.
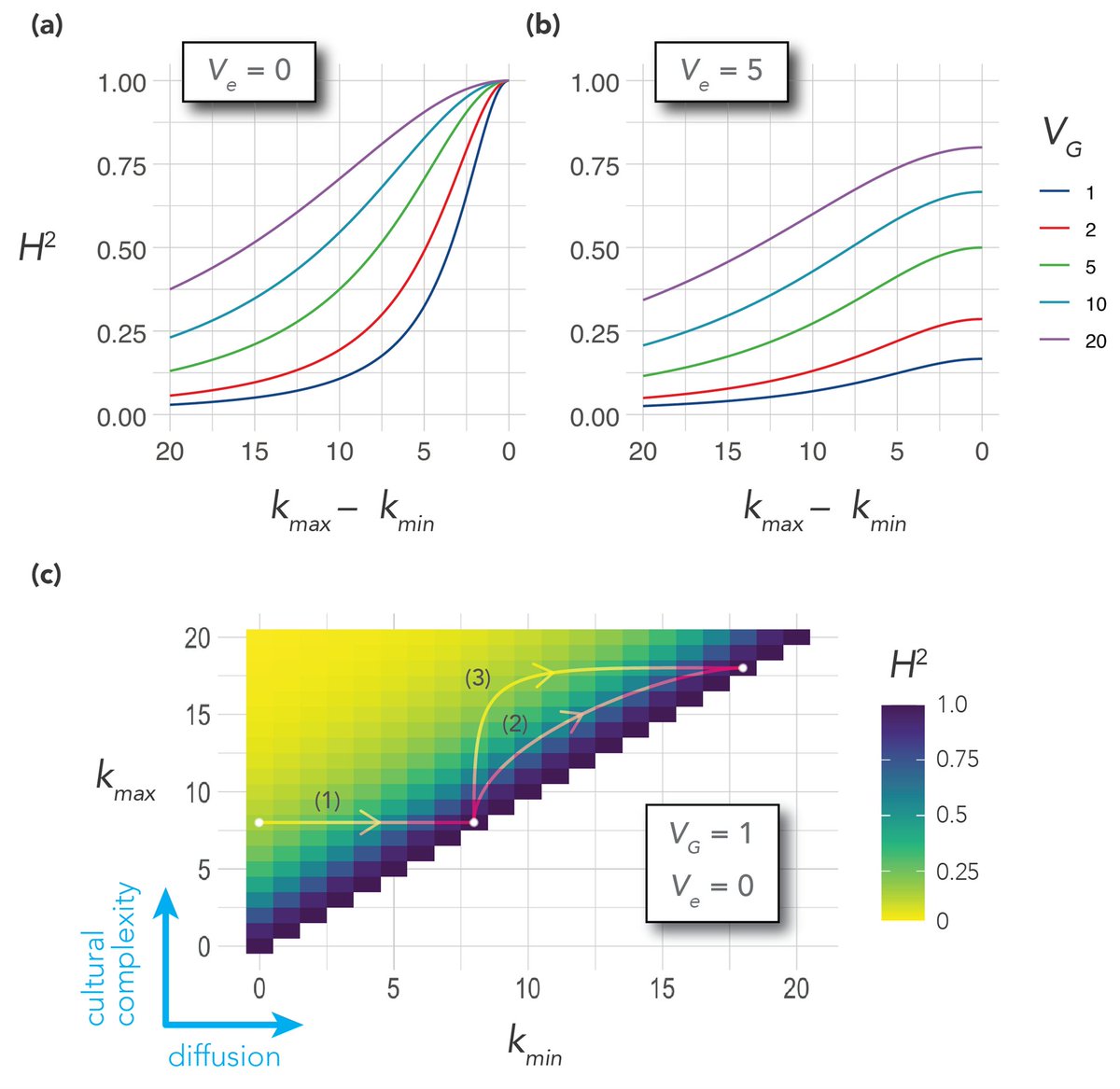
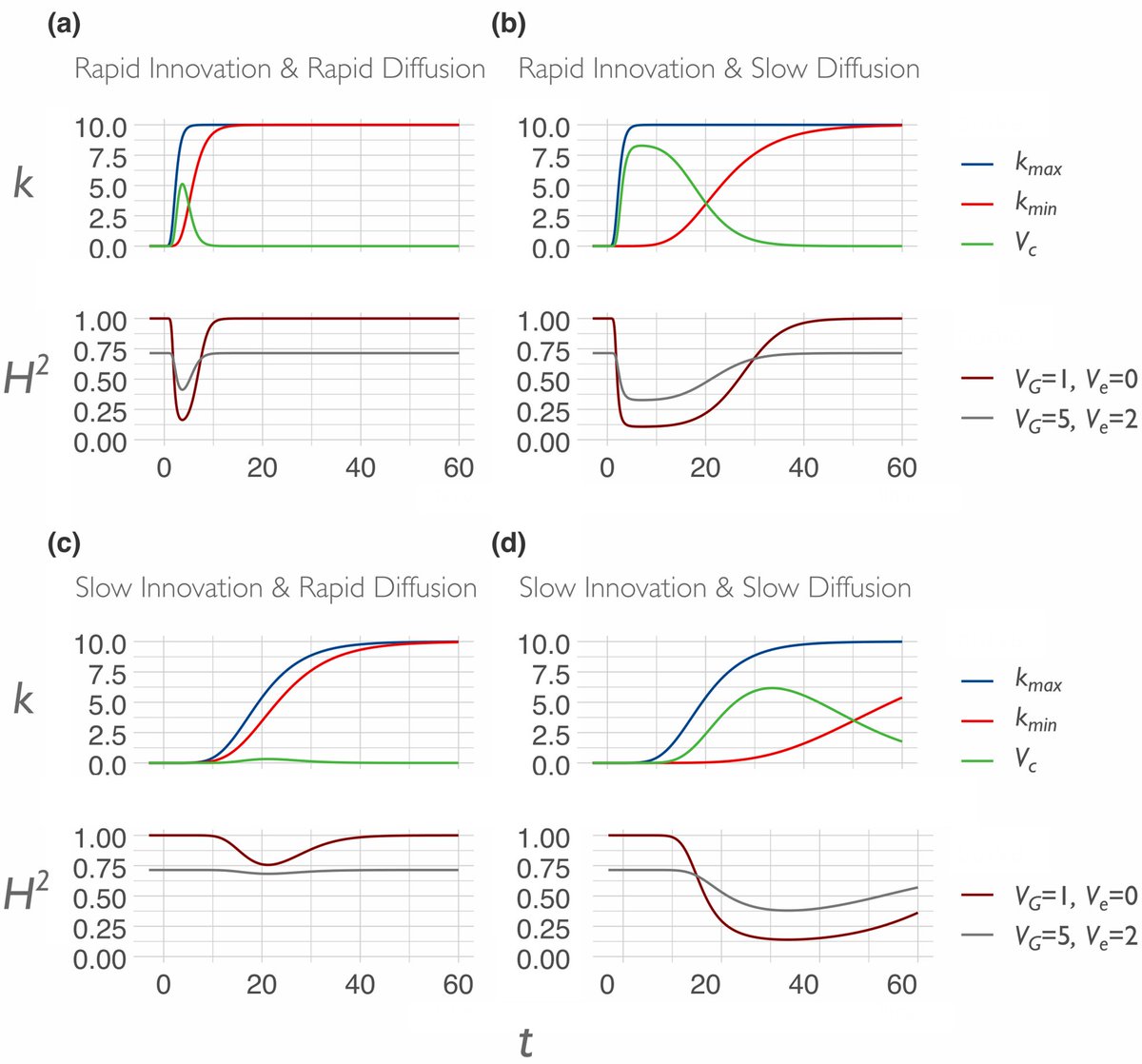
Section 5 and the conclusion tie everything together, but I’ll leave you to read it (muth.io/cegh). There’s a formal model with some pretty graphs in the Appendix:
I presented some in progress theoretical and empirical work on “Hunter-gatherer egalitarianism and the evolution of evil eye beliefs” at the Biological Anthropology seminar series at University College London (UCL) in London, UK.
I gave a general talk on “Cultural Evolution and the Measurement of Culture” at Monash University, Melbourne. I discussed various bits of research including:
Muthukrishna, M. & Henrich, J. (2019). A Problem in Theory. Nature Human Behaviour. [Download]
Muthukrishna, M., Bell, A. V., Henrich, J., Curtin, C., Gedranovich, A., McInerney, J., & Thue, B. (under review). Beyond WEIRD Psychology: Measuring and Mapping Scales of Cultural and Psychological Distance. [Download]
Chudek, M., Muthukrishna, M. & Henrich, J. (2015) Cultural Evolution. In Handbook of Evolutionary Psychology, 2nd Edition. Edited by D. M. Buss. [Download]
Muthukrishna, M., Doebeli, M., Chudek, M., & Henrich, J. (2018). The Cultural Brain Hypothesis: How culture drives brain expansion, sociality, and life history. PLOS Computational Biology, 14(11): e1006504. [Download] [Supplementary]
Stimmler, D. & Muthukrishna, M. (In prep). When Cooperation Promotes Corruption and Undermines Democracy.
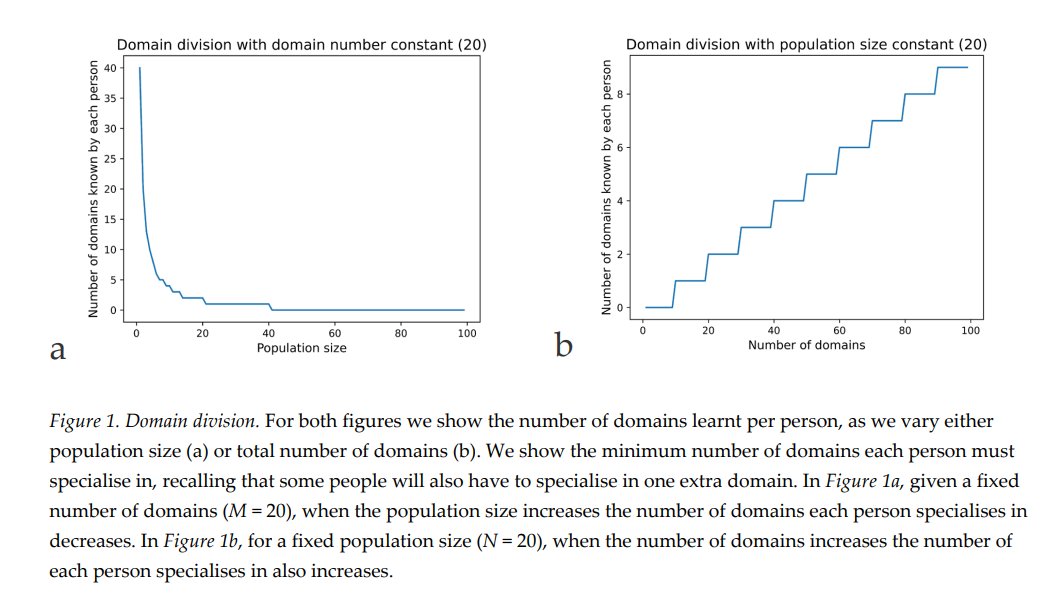
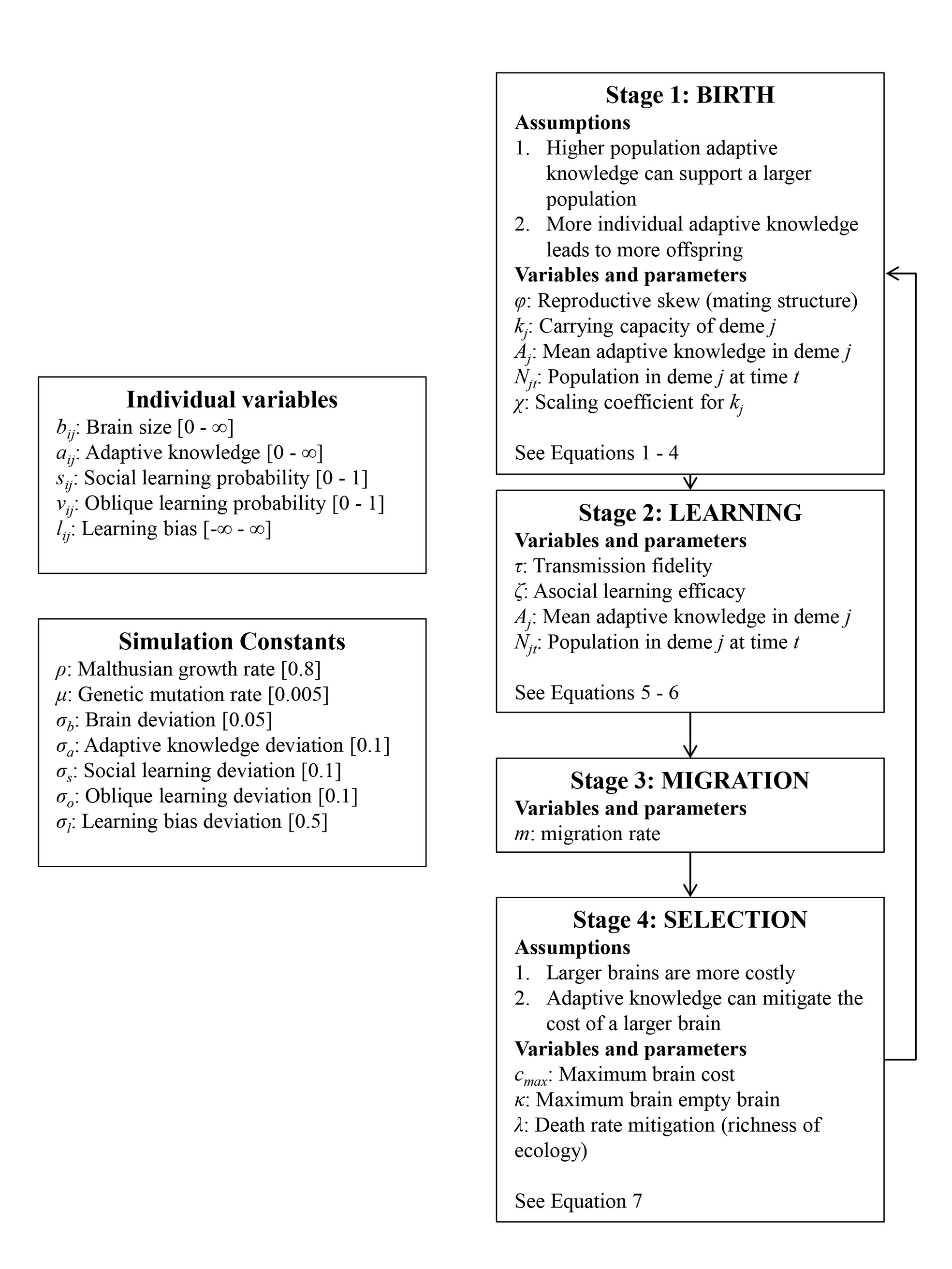
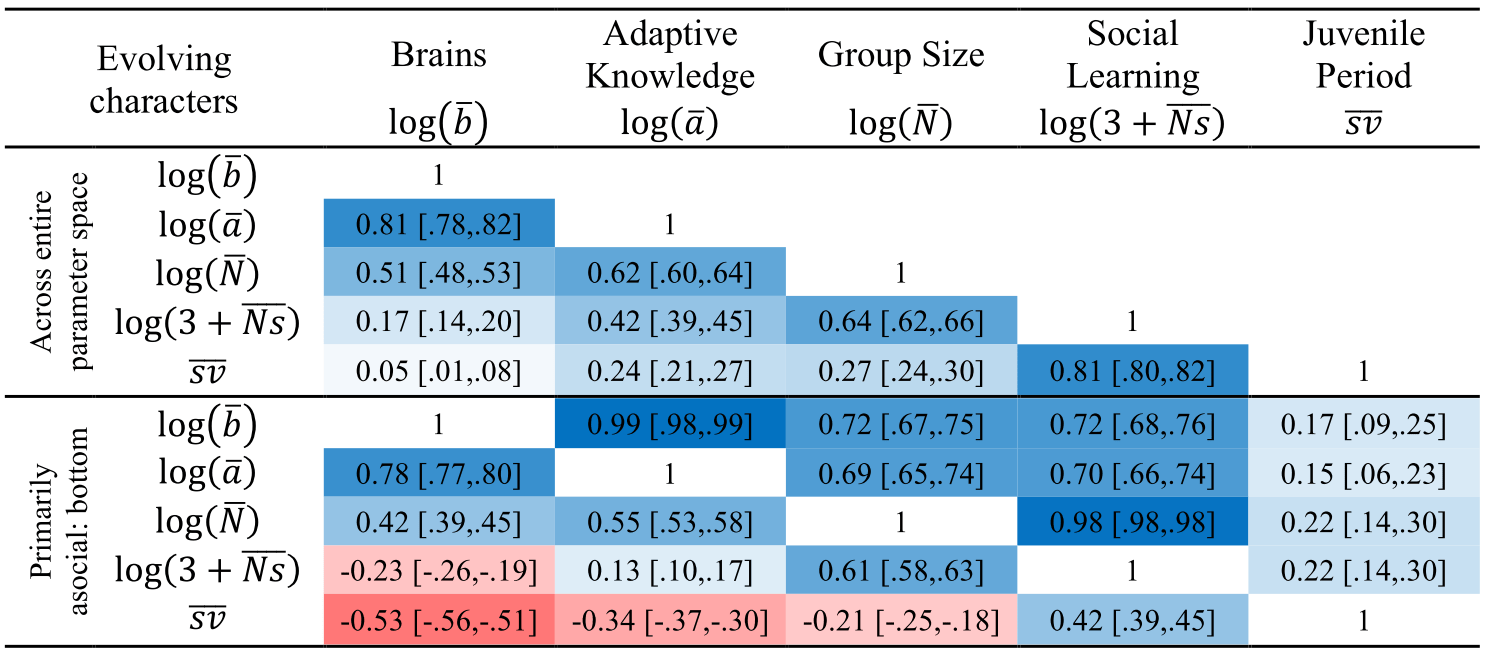
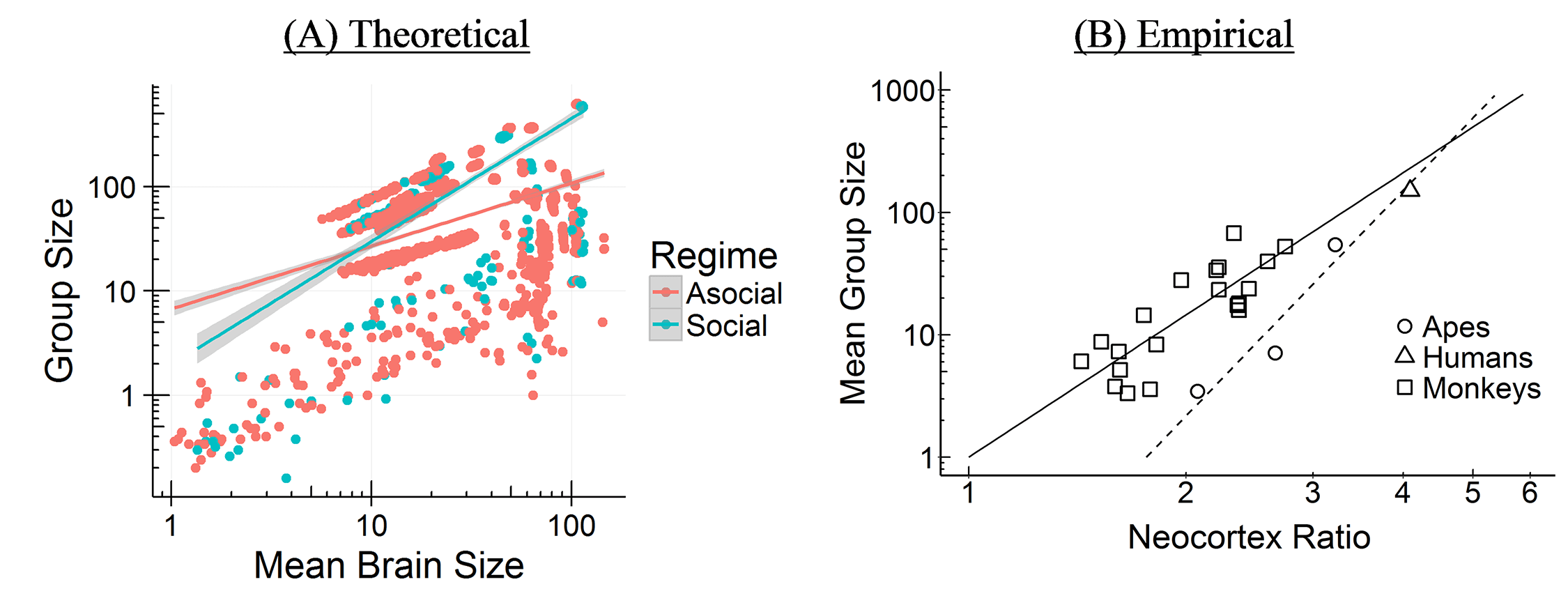
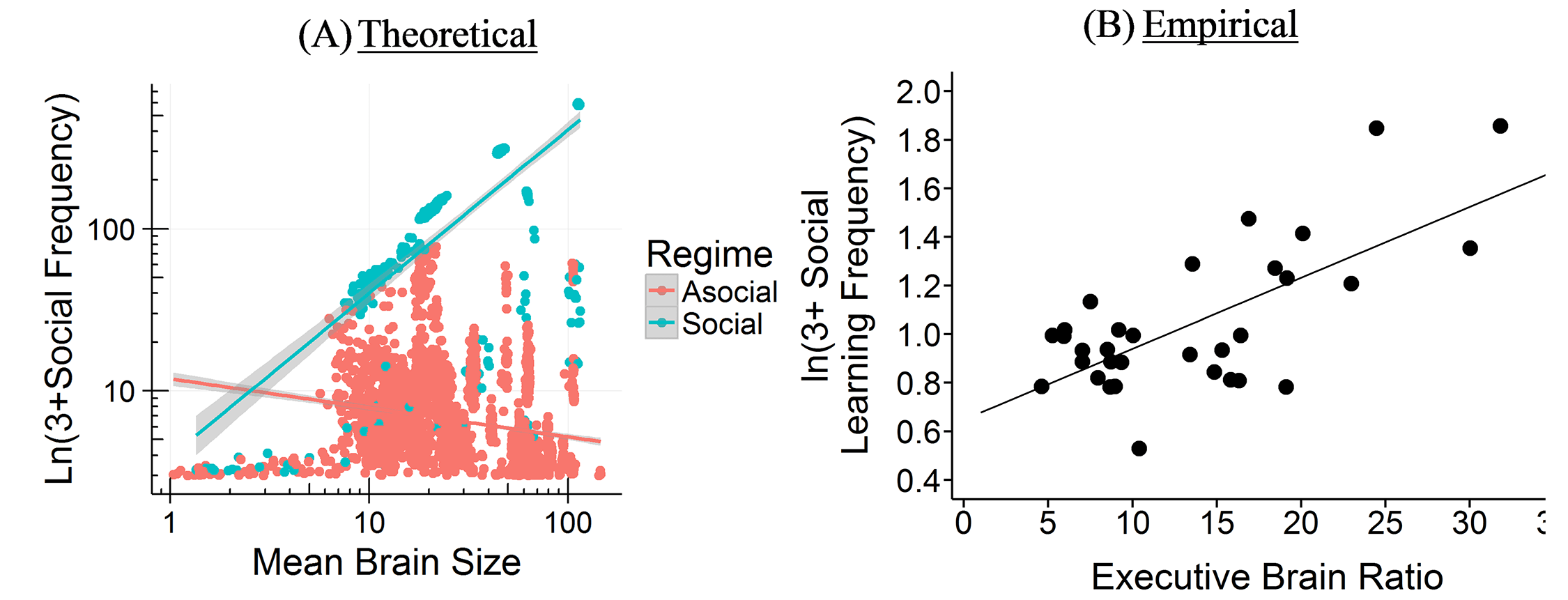
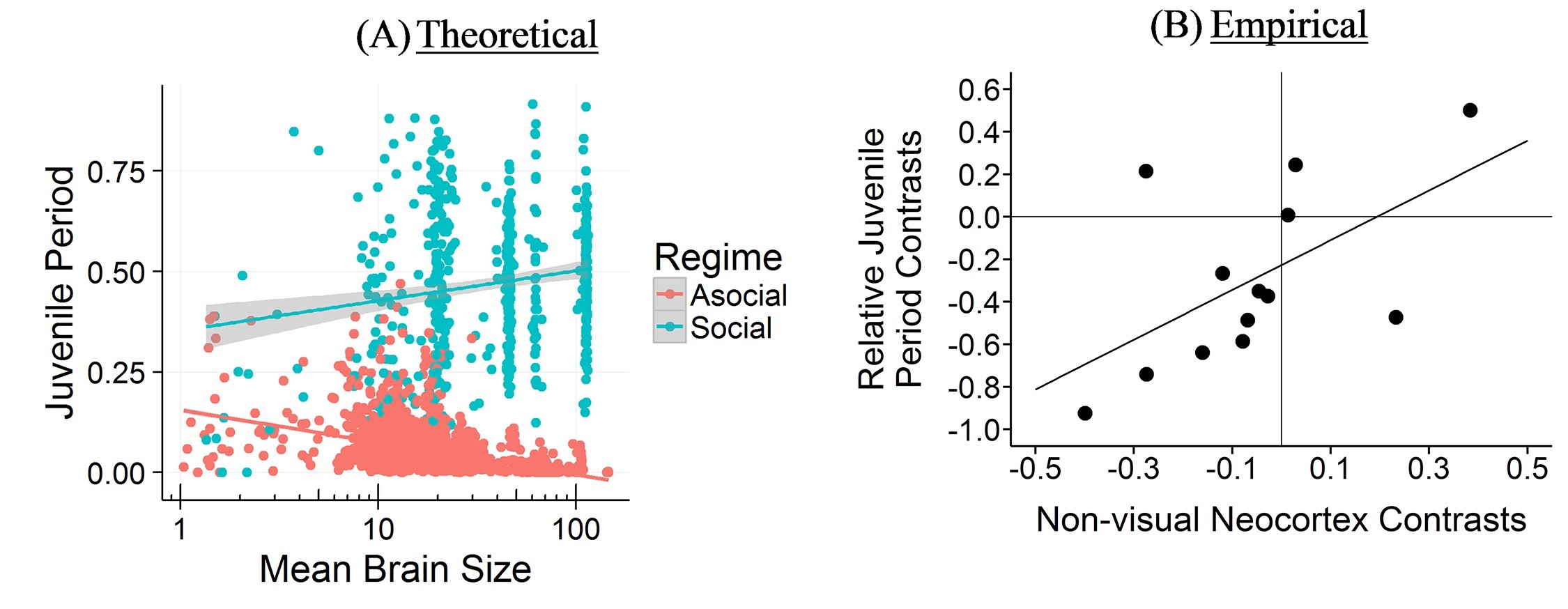
The Cultural Brain Hypothesis is a more general theory for brain evolution across species that unifies more specific explanations around environmental hypotheses and social brain hypotheses. The theory is formalized using an analytical and a computational model.
Figure 1 from paper
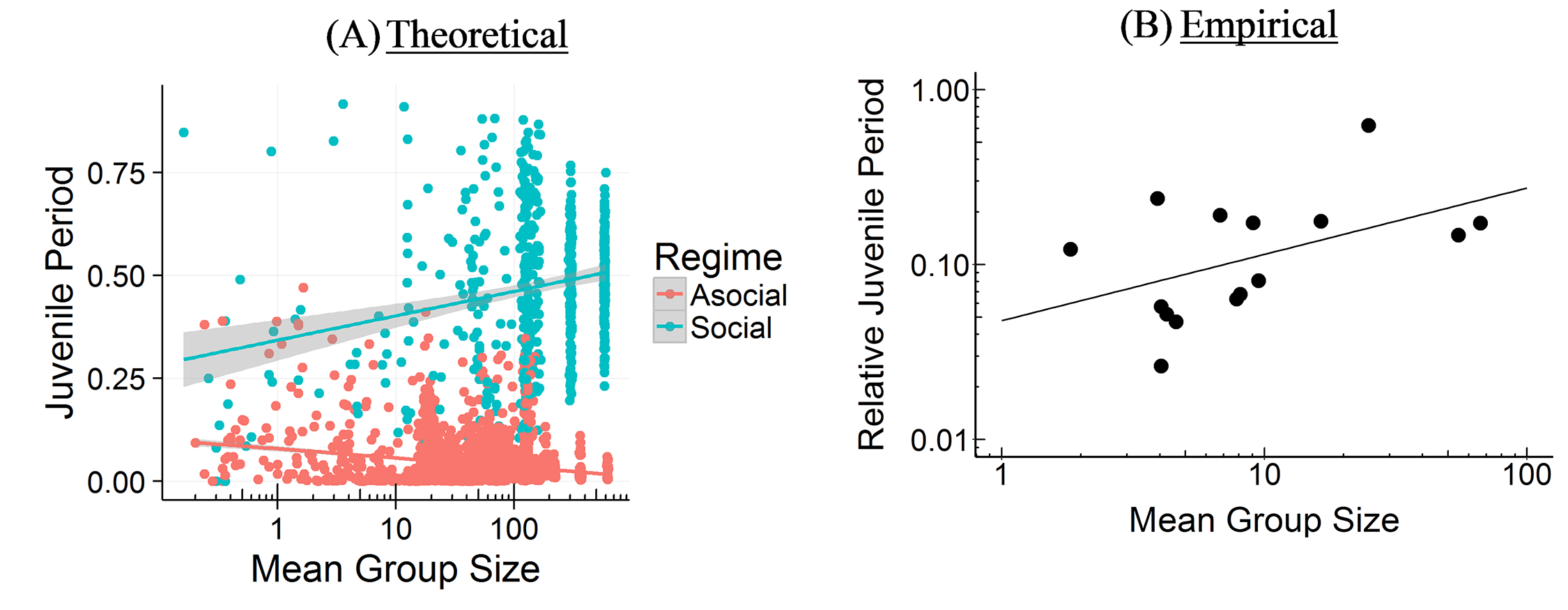
The CBH shows how the environment constrains evolution and how social factors are necessary infrastructure for more social learning species. It predicts different relationships between brain size, sociality, mating structure, the length of the juvenile period, innovation and knowledge, and social learning strategies.
Table 1 from paper.
According to the CBH, the environment constrains brain evolution rather than driving it – brain size is affected by the environment, because you need to have enough calories to feed your brain. But your ability to derive calories from what’s available (or potentially available) is driven by how smart you are – how much information you have. All else being equal, a lush rainforest will have larger brains than an arid desert.
The model specifies two pathways for acquiring this information, both of which can lead to bigger brains – asocial learning and social learning (or some combination of these). If you take the asocial path, you’re reliant on your own intelligence and you don’t have to worry about the social infrastructure. Asocial brains can be larger depending on how easy it is to learn things asocially, but they’ll tend to be smaller than social brains on average.
If you take the social path, it requires all kinds of social infrastructure – more tight-knit and perhaps larger group to learn from, a longer juvenile period, more care during that longer juvenile period, tolerance for other members of the group, an ability and proclivity to learn from other members of the group, and so on. Culture is socially transmitted information, which is a cheaper and more efficient way to get information than asocial learning, but does require all these social factors.
The theory links together ecology and social factors and shows how constraints for learning culture and information in general are what drive the expansion in brain evolution (rather than adaptations to the environment or social factors directly). The model allows us to make sense of a lot of puzzling relationships between brain size, sociality, mating structures, juvenile period, innovation, knowledge, and social learning strategies, and other social and environmental features. We’ve tested some of these relationships among cetaceans and in this paper, we compare it to tests in primates. Unfortunately, most of the focus has been on the more interesting more social learning species (you publish papers by showing how animals and babies are smart and human adults are dumb, not vice versa). The next step is to try to test the predictions for more asocial taxa.
The Cumulative Cultural Brain Hypothesis (CCBH)
The CCBH is a narrow set of parameters that can lead to a take off where information and technology start accumulating faster and faster forcing brains and social factors to evolve to keep up. In our species, our brains continue to grow to the point where we end having trouble giving birth to our babies (larger heads are more difficult to birth), we give birth to our babies prematurely relative to other animals (compare a human infant to a gazelle ready to run). This leads to strategies to take care of our now helpless infants, like forcing fathers to pay for childcare or stick around, and normatively controlling female sexuality so dad knows it’s his. We do other things to keep up. We divide up the information, leading to a division of information and a division of labor (specialization), which can lead to a collective brain. We expand our juvenile period, so we spend longer in childhood, and have an extraordinarily long period of adolescence (the time between when you can reproduce and when you actually do), just to keep learning the ever growing body of information needed to outcompete other members f our group. This last strategy is now at the point we’re hitting a new biological limit – not in the size of the brains we can birth, but in our ability to reproduce at a later age. (I wrote a bit about this for MoneySupermarket in reference to why it takes longer to buy a house).
According to the CCBH, this take off requires:
High transmission fidelity. This could include more cognitive abilities like gaze tracking, shared intentionality, theory of mind, the ability to recognize, distinguish, and imitate potential models, but also more social factors like social tolerance, and ever more sophisticated methods of teaching (consider how long you’ve probably spent in formal education plus internships or low paid entry-level jobs).
Low reproductive skew. Consistent with a “monogamish” or cooperative breeding structure that suppresses reproductive skew. A cooperative breeding environment would have also been ideal to allow for an easy transition to oblique learning. Chimps learn from their mom, but having multiple moms and dads means you can focus on who’s better rather than who you have access to.
Smart ancestors. There is an interaction between transmission fidelity and efficient individual learning. Social learners benefit from smart asocial learners who’s knowledge they can exploit.
Rich ecology. There have to be potential returns in the environment. That is, there are large game or good sources of calories, only requiring the knowledge to acquire them.
There’s more in the paper, which I encourage you to read.
I presented some in progress theoretical and empirical work on “The Evolution of Evil Eye Beliefs and Related Behaviors” at the 2nd Cultural Evolution Society (CES) conference.
Part of this work was based on a recent paper published in Nature Human Behaviour, with some context published in Evonomics and ProMarket (pre-print). But the main part was work in progress on understanding the evolution of evil eye beliefs and hunter-gatherer egalitarianism.
I spent the last few days as a meeting organized by the Templeton World Charity Foundation (TWCF) at the Diverse Intelligences Summer Institute Meeting at University of St Andrews, St Andrews, Scotland. It was an amazing few days of stimulating conversation on intelligence.
I also discussed the general approach to understanding corruption using cultural evolution and the science of cooperation – corruption is one scale of cooperation undermining another. For example, nepotism is cooperation at the scale of kin, well explained by inclusive fitness, undermining cooperation at the scale of the formal institution. More on this framework can be found at ProMarket or Evonomics. Finally, I presented some work in progress based on this approach, including some work by my students.
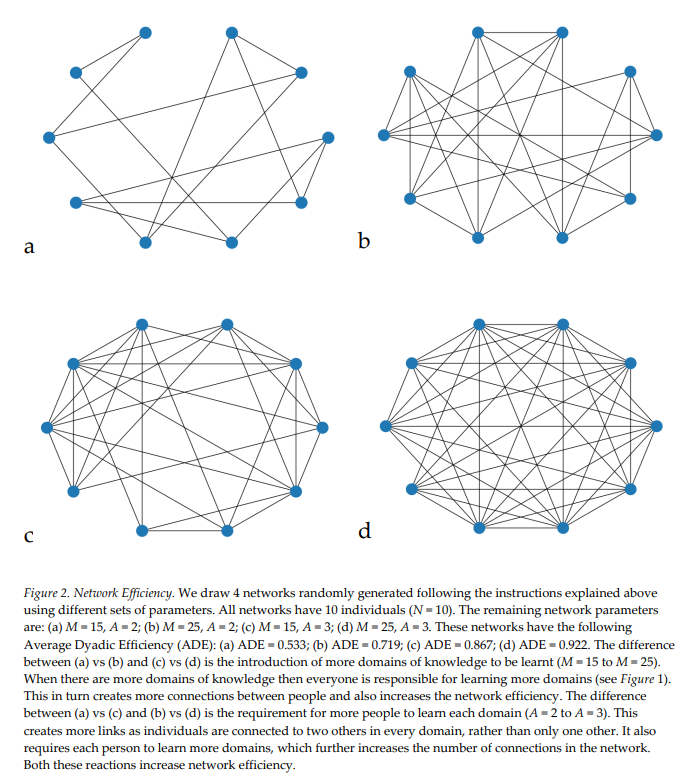
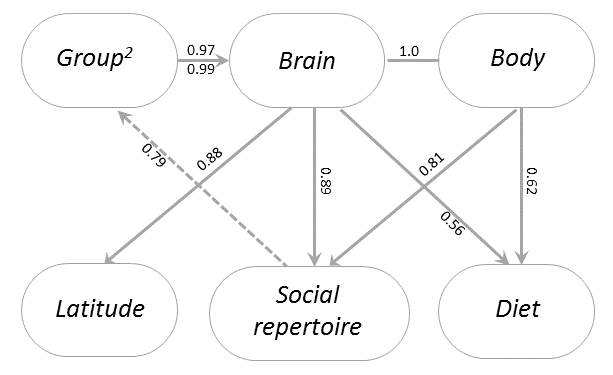
Last week, my paper with Kieran Fox and Susanne Shultz was published in Nature Ecology and Evolution. The paper was a multiyear project, which consisted of countless hours spent poring through marine mammal literature to create the most comprehensive database of cetacean physiology, social structure, life history, and behavior to date. We then used this database to test some of the predictions of the Social Brain and Cultural Brain Hypotheses. Some of the confirmations of these predictions are shown in Figure 3 of the paper below.
Cetaceans represent a great test for the Social Brain and Cultural Brain Hypotheses (CBH), because of how evolutionarily alien these species are, and how strange their underwater world is compared to the world we inhabit. We have previously tested the CBH predictions with primates, but their evolutionary closeness to humans means that the relationships we find may be due to our evolutionary logic or due to these features (such as large brains and high sociality) being present in a common ancestor. Thus finding these relationships in cetaceans is strong evidence for the evolutionary logic. It also sets up cetaceans as an interesting control group for understanding human evolution.
The ongoing massive media response and public interest in marine mammals and the evolutionary sciences was heartwarming. Altmetrics suggested a score of 1026, in the top 5 of articles in Nature Ecology and Evolution, receiving the most attention of recent articles and top 50 of all articles of a similar age. Highlights included several video and audio interviews, including with BBC World News, BBC World Service Radio “Science in Action”, CBC “The Broadcast” (below), and the front page of the print edition of the The Times and front page of the website of The Guardian.
Introducing the possibility of bribes into an institutional punishment public goods game results in reduced contributions.
In an institutional punishment public goods game, stronger leaders result in more cooperation. In our modified “bribery game”, stronger leaders result in less cooperation.
Anti-corruption measures including transparency and tying leaders payoffs to the success of the public good result improve contributions, except if economic potential is low and leaders are weak. Here, they can actually further reduce contributions.
Culture matters. Exposure to corrupt norms via living in corrupt places increases bribes, but having an ethnic heritage that includes corrupt countries, but not having actually lived there yourself results in less bribery.
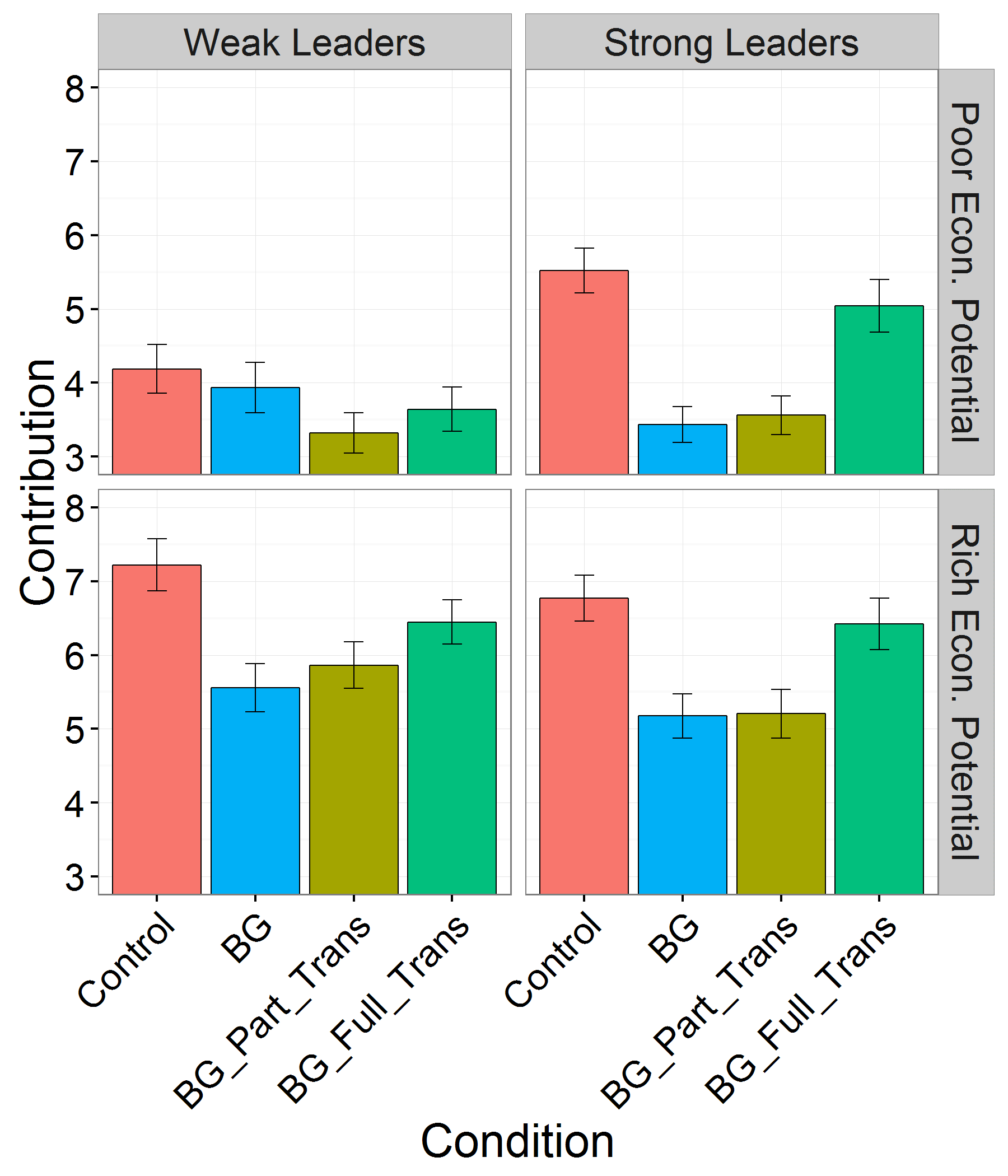
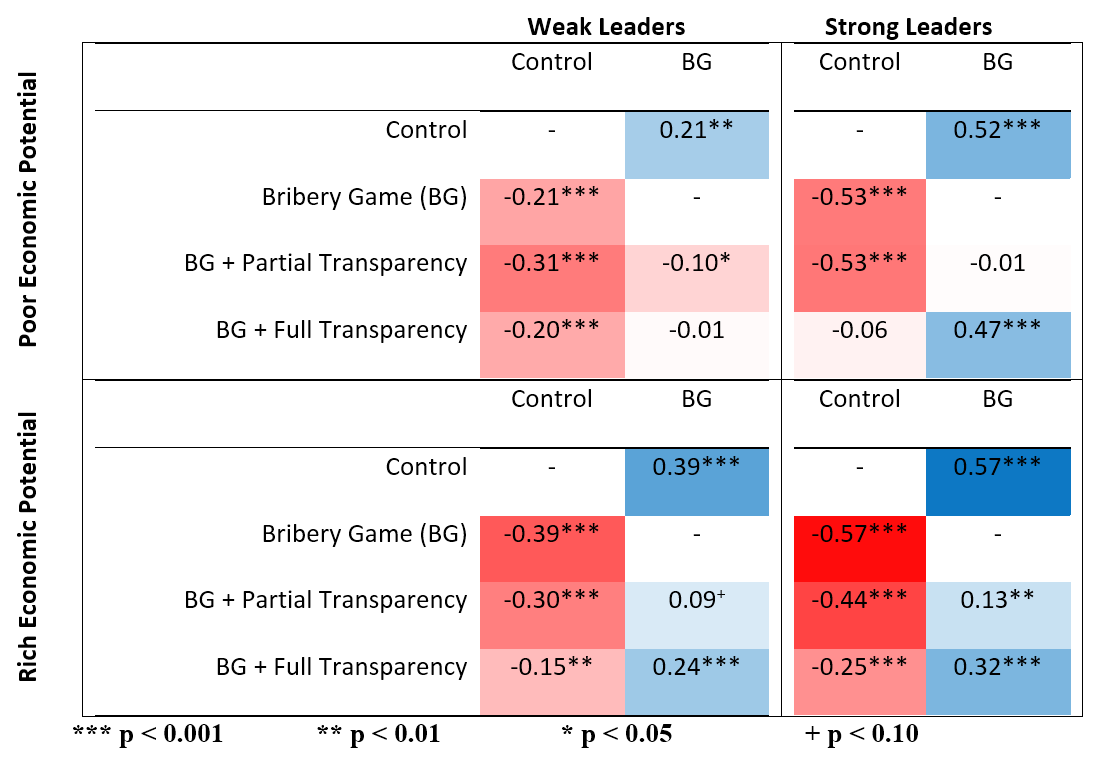
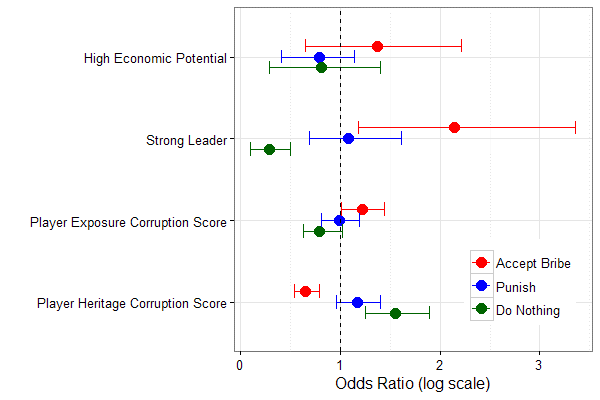
Figures 1, 2 and 3, reproduced below illustrate these results.
Raw contributions (of the ten endowed points) and 95% confidence intervals for each within-subject treatment (control, BG, BG with partial transparency or BG with full transparency) in each between-subjects structural context (strong versus weak leader and poor versus rich economic potential). These data are consistent with our theory that predicts that more powerful leaders increase contributions in the IPGG but decrease contributions in the BG.
Darker blue indicates greater public goods provisioning and darker red indicates reduced public goods provisioning. All coefficients were extracted from a single model by changing reference groups. The columns represent the reference group treatment (control versus BG), while each row shows the coefficient of each treatment compared with this reference group. The contributions were z scores, so the coefficients represent s.d. The full model is reported in the Supplementary Information. In all models, we accounted for the clustering inherent in the experimental design by including a fixed effect for the number of subjects and random effects for participants within groups. Note that in all treatments and structural contexts, the BG has lower contributions than the structurally equivalent IPGG (control). Corruption mitigation effectively increases contributions (although not to control levels) when leaders are strong or the economic potential is rich. When leaders are weak and the economic potential is poor, the apparent corruption mitigation strategy, full transparency has no effect and partial transparency further decreases contributions. *P < 0.10; **P < 0.05; ***P < 0.01; ****P < 0.001.
Odds ratios and 95% confidence intervals are shown for each behaviour (accept bribe, punish or do nothing).
There is nothing natural1 about democracy. There is nothing natural about living in communities with complete strangers. There is nothing natural about large-scale anonymous cooperation. Yet, this morning, I bought a coffee from Starbucks with no fear of being poisoned or cheated. I caught a train on London’s underground packed with people I’ve never met before and will probably never meet again. If we were commuting chimps in a space that small, it would have been a scene out of the latest Planet of the Apes by the time we reached Holborn station. We’ll return to this mystery in a moment.
There is something very natural about prioritizing your family over other people. There is something very natural about helping your friends and others in your social circle. And there is something very natural about returning favors given to you. These are all smaller scales of cooperation that we share with other animals and that are well described by the math of evolutionary biology. The trouble is that these smaller scales of cooperation can undermine the larger-scale cooperation of modern states. Although corruption is often thought of as a falling from grace, a challenge to the normal functioning state—it’s in the etymology of the word—it’s perhaps better understood as the flip side of cooperation. One scale of cooperation, typically the one that’s smaller and easier to sustain, undermines another.
When a leader gives his daughter a government contract, it’s nepotism. But it’s also cooperation at the level of the family, well explained by inclusive fitness2, undermining cooperation at the level of the state. When a manager gives her friend a job, it’s cronyism. But it’s also cooperation at the level of friends, well explained by reciprocal altruism3, undermining the meritocracy. Bribery is a cooperative act between two people, and so on. It’s no surprise that family-oriented cultures like India and China are also high on corruption, particularly nepotism. Even in the Western world, it’s no surprise that Australia, a country of mates, might be susceptible to cronyism. Or that breaking down kin networks predicts lower corruption and more successful democracies (Akbari, Bahrami-Rad & Kimbrough, 2017; Schulz, 2017). Part of the problem is that these smaller scales of cooperation are easier to sustain and explain than the kind of large-scale anonymous cooperation that we in the Western world have grown accustomed to.
So how is it that some states prevent these smaller scales of cooperation from undermining large-scale anonymous cooperation? The typical answer is that more successful nations have better institutions. All that’s required is the right set of rules to make society function. But even on the face of it, this answer seems incomplete. If it were true, Liberia, who borrowed more than its flag from the United States, ought to be much more successful than it is4. Instead, these institutions are supported by invisible cultural pillars without which the institutions would fail. For example, without a belief in rule of law—that the law applies to all and cannot be changed on the whim of the leader—it doesn’t matter what the constitution or legal code says, no one is listening. Without a long time horizon, decisions are judged on how well they serve our immediate needs making larger-scale projects, like reducing the effects of Climate Change, harder to justify5. Similarly, institutions often lack the punitive power to actually punish perpetrators. For example, most people in the US and UK pay their taxes, even though in reality the IRS and Her Majesty’s Revenue and Customs lack the power to prosecute widespread non-compliance; your probability of getting caught is low. The tax compliant majority may never discover that they can cheat or how to get away with it (Chetty, et al. 2013) and they may not actively seek this information as long as the probability of getting caught is non-zero, the system seems fair, and it seems like everyone else is complying. Or in other words, it’s a combination of norms and institutions. But, it gets tricky—institutions are themselves hardened or codified norms6 and the norms themselves evolve in response to the present environment and due to path-dependence of previous environments, past decisions, and the places migrants come from. Modern groups vary on individualism (Talhelm, et al., 2014) and even sexist attitudes (Alesina, et al., 2013) based on their ancestors’ farming practices7. The science of cultural evolution describes the evolution of these norms and introduces the possibility of out-of-equilibria behavior (people behaving in ways that do not benefit them individually) for long enough for institutions to try to stabilize the new equilibria. For a summary of cultural evolution, see Joseph Henrich’s excellent book and for an even shorter summary see this chapter). How do we begin to understand these processes?
The real world is messy and before we start running randomized control trials or preparing case studies, it’s useful to model the basic dynamics of cooperation using a simpler form that gets at the core elements of the challenge. One commonly used model is called the “Public Goods Game”. The gist of the game is that I give you, and say 9 others, $10. Whatever you put into a pool (the public good), I’ll multiply by say 3, but then I’ll divide the money equally regardless of contribution. This is similar to paying your taxes for public goods that we all benefit from, like roads, clean water, or environmental protections. The dilemma is this: the best move is for everyone to put all their money in the pool. Then they’ll all go home with $30. But it’s in my best interests to put nothing in the pool and let everyone else put their money in. If I put in nothing and they put in $10 each, I’ll go home with almost $40 ($10*9*3people / 10 = $37). What happens when we play this game?
Well, if we play it in a WEIRD8 nation, where prosocial norms tend to be higher, people put about half their money in, but as they gradually realize they can make more by putting in less, contributions dwindle to zero. One way to sustain contributions is to introduce peer punishment—allow people to spend some portion of their money to punish other people. This is similar to the kind of punishment we might see in a small village. I know who you are or at least I know your parents or people you know. If you steal my crops, I’ll punish you myself or ruin your reputation. In the game, if we introduce the possibility of peer punishment, contributions rise again. The problem is that this doesn’t scale well. As the number of people grows, we get second-order free-riding—people prefer to let someone else pay the cost of punishment. When someone cuts a queue, you grumble—someone ought to tell that person off! Someone other than me… And you can also get counter-punishment—revenge for being punished. The best solution seems to be to create a punishment institution. Pick one person as a “Leader” and allow them to extract taxes that can be used to punish free-riders. This works really well and scales up nicely. It’s similar to a functioning police force and judiciary in WEIRD nations. In fact, the models suggest that the more power you give to the leader, the more cooperation they can sustain. Aha! Problem solved. Not quite. Models like these are very useful for distilling the core of a phenomenon, they can miss things. Recall where we started—smaller-scales of cooperation can undermine the larger-scale.
In our recently published paper, we wanted to show just how easy it was to break that well-functioning institution. We did it by introducing the possibility of another very simple form of cooperation—you scratch my back, I’ll scratch yours—bribery. And then we wanted to show the power of invisible cultural pillars by measuring people’s cultural background and by trying to fix corruption using common anti-corruption strategies. We wanted to show that these strategies, including transparency, don’t work in all contexts and can even backfire.
Our “Bribery Game” was the usual institutional punishment public goods game with the punishing leader, but with one additional choice—players could not only keep money for themselves or contribute to the public pool, they could also contribute to the leader. And the leader could not only punish or not punish, they could instead accept that contribution. What happened? On average, we saw contributions fall by 25% compared to the game without bribery as an option. More than double what the pound has fallen against the USD since Brexit (~12%9). Fine, bribery is costly. The World Bank estimates $1 trillion is paid in bribes alone; in Kenya, 8 out of 10 interactions with public officials involves a bribe, and as Manfred Milinski points out in his summary of our paper, most of humanity—6 billion people—live in nations with high levels of corruption. Our model also reveals that unlike the typical institutional punishment public goods game, where stronger institutions mean that more cooperation can be sustained, when bribery is an option, stronger institutions mean more bribery. A small bribe multiplied by the number of players will make you a lot richer than your share of the public good! So can we fix it?
The usual answer is transparency. There are also some interesting approaches, like tying a leader’s salary to the country’s GDP—the Singaporean model10. So what happened when we introduced these strategies? Well, when the public goods multiplier was high (economic potential—potential to make money using legitimate means—was high) or the institution had power to punish, then contributions went up. Not to levels without bribery as an option, but higher. But in poor contexts with weak punishing institutions, transparency had no effect or backfired. As did the Singaporean model11. Why? Consider what transparency does. It tells us what people are doing. But as psychological and cultural evolutionary research reveals, this solves a common knowledge problem and reveals the descriptive norm—what people are doing. For it to have any hope of changing behavior, we need a prescriptive or proscriptive norm against corruption. Without this, transparency just reinforces that everyone is accepting bribes and you’d be a fool not to. People who have lived in corrupt countries will have felt this frustration first hand. There’s a sense that it’s not about bad apples—the society is broken in ways that are sometimes difficult to articulate. But societal norms are not arbitrary. They are adapted to the local environment and influenced by historical contexts. In our experiment, the parameters created the environment. If there really is no easy way to legitimately make money and the state doesn’t have the power to punish free-riders, then bribery really is the right option. So even among Canadians, admittedly some of the nicest people in the world, in these in-game parameters, corruption was difficult to eradicate. When the country is poor and the state has no power, transparency doesn’t tell you not to pay a bribe, it solves a different problem—it tells you the price of the bribe. Not “should I pay”, but “how much”?
There were some other nuances to the experiment that deserve follow up. If we had played the game in Cameroon instead of Canada, we suspect baseline bribery would have been higher. Indeed, people with direct exposure to corruption norms encouraged more corruption in the game controlling for ethnic background. And those with an ethnic background that included more corrupt countries, but without direct exposure were actually better cooperators than the 3rd generation+ Canadians. These results may reveal some of the effects of migration and historical path dependence. Of course, great caution is required in applying these results to the messiness of the real world. We hope to further investigate these cultural patterns in future work. The experiment also reveals that corruption may be quite high in developed countries, but its costs aren’t as easily felt. Leaders in richer nations like the United States may accept “bribes” in the form of lobbying or campaign funding and these may indeed be costly for the efficiency of the economy, but it may be the difference between a city building 25 or 20 schools. In a poor country similar corruption may be the difference between a city building 3 or 1 school. Five is more than 3, but 3 is three times more than 1. In a rich nation, the cost of corruption may be larger in absolute value, but in a poorer nation, it may be larger in relative value and felt more acutely.
The take home is that cooperation and corruption are two sides of the same coin; different scales of cooperation competing. This approach gives us a powerful theoretical and empirical toolkit for developing a framework for understanding corruption, why some states succeed and others fail, why some oscillate, and the triggers that may lead to failed states succeeding and successful states failing. Our cultural evolutionary biases lead us to look for whom to learn from and perhaps whom to avoid. They lead us to blame individuals for corruption. But just as atrocities are the acts of many humans cooperating toward an evil end, corruption is a feature of a society not individuals. Indeed, corruption is arguably easier to understand than my fearless acceptance of my anonymous barista’s coffee. Our tendency to favor those who share copies of our genes—a tendency all animals share—lead to both love of family and nepotism. Putting our buddies before others is as ancient as our species, but it creates inefficiencies in a meritocracy. Innovations are often the result of applying well-established approaches in one area to the problems of another. We hope the science of cooperation and cultural evolution will give us new tools in combating corruption.
1 Putting aside what it means for something to be natural for our species, suffice to say these are recent inventions in our evolutionary history, by no means culturally universal, and not shared by our closest cousins.
2 Genes that identify and favor copies of themselves will spread.
4 The United Nations Human Development Index ranks the United States 10th in the world. Liberia is 177th.
5Temporal discounting the degree to which we value the future less than the present. Our tendency to value the present over the future is one reason we don’t yet have Moon or Mars colonies, but the degree to which we do this varies from society to society.
6 Written laws can serve a signaling and coordination function; rather than having to interpret norms from the environment. When previously contentious norms are sufficiently well established, you may do well to codify them in law (legislating before they are established might mean more punishment—consider the history of prohibition in the United States).
7 Not that agriculture is the main reason for these cultural differences!
10 Singapore’s leaders are the highest paid in the world, but the nation also has one of the lowest corruption rates in the world—lower than the Netherlands, Canada, Germany, UK, Australia, and United States [source].
11 Note, there are some conceptual issues that make interpretation of the Singaporean treatment ambiguous. We discuss this in the supplementary. We’ll have to further explore this in a future study. Such is science.