Considering the limitations of psychology today, I discussed ways in which we can move beyond WEIRD (Western, Educated, Industrialized, Rich, and Democratic) psychology in a global collaborative manner and strengthen the links between basic and applied policy research. I also discussed the importance of historical psychology.
Many thanks to Joseph Henrich for the enthusiastic introduction.
Long post, but an important topic that helps to resolve controversies such as IQ differences.
Quick summary: We reconcile behavioral genetics and cultural evolution under a dual inheritance framework. A cultural evolutionary behavioral genetic approach cuts through the nature–nurture debate and helps resolve controversies such as IQ.
Business is booming in behavioral genetics. We’re in the midst of a GWAS gold rush. Powerful computers and sequenced DNA of millions has led to an industrious search for SNPs that correlate with a variety of traits. Some even claim the curse of reverse causality has been lifted.
There’s also been a parallel revolution in cultural psychology and cultural evolution. Genes, culture, and the environment have often co-evolved, shaping our species. But the revolutions in behavioral genetics and cultural evolution have largely been independent. But, given the extensiveness of the cultural and culturally-shaped environment, cultural evolution offers an important but typically missing complement to otherwise insightful methodological and empirical analyses within behavioral genetics. Genes and culture are intertwined. For example, our jaws too weak and guts too short for a world w/o controlled fire & cooked food. It’s obvious that lower environmental variation will lead to higher heritability scores. Less obvious is how culture can mask or unmask genetic variation. Or how diffusion and innovation can increase or decrease heritability. Or how to define a single society for the purposes of measuring heritability, without being able to identify cultural cleavages that can lead to Scarr-Rowe type effects: en.wikipedia.org/wiki/Scarr-Row…
Reconciling behavioral genetics & cultural evolution offers insights for differences in heritability between and within populations, differences in heritability across development, and the rise in IQ (Flynn Effect). We’d like a discussion that nuances common interpretations of the nature and nurture of behavior.
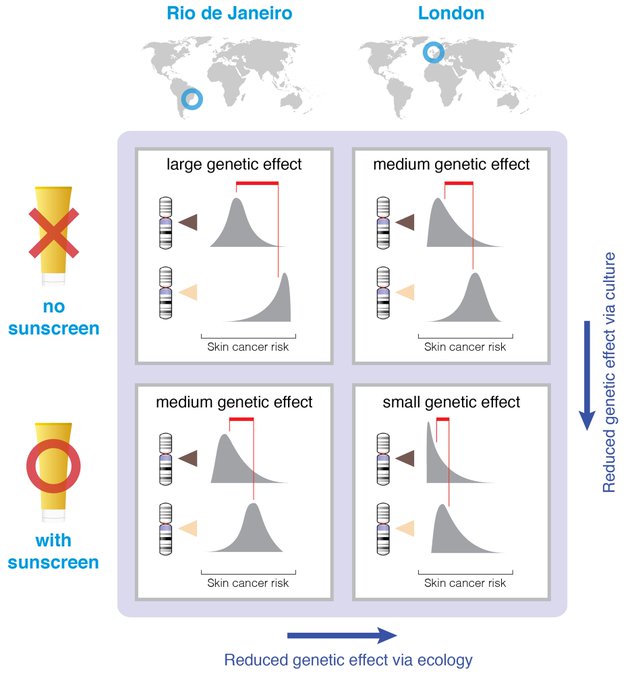
First, let’s quickly get a common misinterpretation of heritability out of the way: Heritability is not an index of the genetic basis of a trait nor a measure of the relative contribution of nature compared to nurture. It’s the proportion of phenotypic variance for some trait that is explained by genetic variance. So obviously variability in genes, in environment, and in traits will all matter. A quick illustration: skin pigmentation and UV levels. Genes affect level of skin pigmentation and propensity for tanning instead of burning; ancestral adaptations to UV radiation at different latitudes.
Migration means melanin is mismatched to latitude: Aussies with European ancestry are more susceptible to skin cancer; Europeans with African and South Asian ancestry have higher rates of vitamin D deficiency.
A Gene×Environment approach won’t predict how heritability estimates change over time as non-genetic adaptations compensate for genetic mismatches: fairer Australians wear sunscreen, a hat, & covered clothing; darker Europeans consume vitamin D supplements & vitamind D-rich or fortified foods. Here it’s easier to see that heritability is a function not only of genes, traits, and ecology, but also of an evolving cultural environment. The environment is not an inert backdrop against which genes should be evaluated. It evolves in relation to both genes and ecology.
Four lessons before we continue. The first is obvious to behavioral geneticists, the second sometimes noted, the third and fourth are typically missing.
Lesson 1
There is no overarching, one-quantity heritability of a trait to be discovered. There is no fixed answer to the question, “What is the heritability of skin cancer?”
Lesson 2
Heritability will depend not only on ecology, but also on culture & specifically on diffusion and innovation—both of which can rapidly change and therefore rapidly change heritability estimates.
Lesson 3
Diffusion and innovation are broadly directional. Cultural diffusion of sunscreen, clothing, shade & sunglasses, and cultural innovation toward more effective screening and treatment of melanomas all work to reduce heritability estimates due to a masking effect.
Were any of these an example of culture unmasking genetic effects, such as tanning salons that induce differential risk according to skin pigmentation level, we would have predicted an increase in heritability.
Lesson 4
We might expect a stronger cultural response where ecological and cultural selection pressures are stronger—skin cancer mitigation in Australia but Vitamin D supplementation in northern Europe. Not been tested to our knowledge, but the predictions are clear.
“Which SNPs are associated with skin cancer?” is similarly culturally dependent. Societies where sunscreen use is common, we expect SNPs associated with skin pigmentation to be less predictive of skin cancer compared to societies where this is not the case.
Similarly, we would expect SNPs associated with antioxidant metabolism to be less predictive of skin cancer in societies whose foods are rich in antioxidants—such as in traditional Mediterranean cuisine.
Section 2.2 is on how cultural evolution shapes heritability through diffusion and innovation. We live on the peaks climbed by cultural evolution – human environments have already been shaped by cumulative cultural evolution—functionally overlapping with genetic evolution.
Diffusion and invention can mask or unmask genes. Examples using language, fertility, and schooling.
Language example
If Cantonese or Yoruba (both tonal) spread, heritability of language ability would increase proportional to variation in “tone” genes.
If Norwegian or Russian (both non-tonal) spread in the same population, heritability of language ability would decrease.
Fertility example
Contraception and social values in 20th century unmasked the effect of genes associated with reproductive behaviors and preferences (heritability rose in US). But a one-child policy or rigid childbearing norms masks the genetic effect.
School example
School is a powerful mechanism for cultural diffusion. Heritability of literacy in:
Australia: Kindergarten: 0.84 Grade 1: 0.80
Scandinavia: Kindergarten: 0.33 Grade: 0.79
Why?
Cultural diffusion of literacy.
Australian children begin receiving compulsory literacy instruction in kindergarten, while in Scandinavia the kindergarten curriculum emphasizes social, emotional, and aesthetic development—literacy instruction only begins in Grade 1.
Assessing the genetic basis of literacy without accounting for particulars of curricula on cultural diffusion is a selection bias of unknown magnitude. Note that literacy in the home environment is already shaped by cultural evolution; there is no ‘baseline’ heritability. Heritability is a composite measure that captures both genes and culture. Saying literacy heritability in Scandinavia jumps up to 0.79 in Grade 1 reveals as much if not more about the disseminative power of modern schooling than it does about the genetic basis of literacy. Similar dynamics with innovation. Read about it in the paper.
However, one neglected factor is “cultural clustering”, where even highly useful forms of cultural knowledge may not easily permeate social barriers. Not necessarily ethnic boundaries, also class, wealth, occupation, political alignment, religion, or incidental geographic layout. Greater differential clustering can lead to a cultural Simpson’s paradox (we’ll get to that shortly, but see Section 3.4).
Cultural FST (CFST) is useful for identifying these clusters. You can read more about that paper here: https://www.michael.muthukrishna.com/beyond-weird-psychology-measuring-and-mapping-scales-of-cultural-and-psychological-distance/
Hopefully, you can see the importance of a cultural evolutionary behavioral genetics. We hope this target article will spark a vibrant discussion. But let’s move onto the problems that obscure the effect of culture:
(1) the WEIRD Sampling Problem
(2) the Hidden Cluster Problem
(3) the Causal Locus Problem
And then describe the:
(4) Cultural Simpson’s Paradox that emerges at their junction.
The WEIRD Sampling Problem
The WEIRD people problem? Pretty bad in genetics too. Twin studies: 94% Western: 60% US, UK, Aus; 25% Nordic 6% Non-Western: 4% China, Japan, South Korea, Taiwan
Remainder of the world, i.e. vast majority of humans are the remaining 2%
Same story in GWAS: 88% European ancestry. 72% from just 3 countries: US, UK, & Iceland 20% from Japan, China, and South Korea
From a cultural evolutionary perspective, given (a) cultural environment, (b) coevolution b/w culture & genes, & (c) cultural differences between populations, not surprising that: 1.Polygenic scores don’t translate well across ancestry groups (European scores, 42% in Africa) 2. Polygenic scores are highly sensitive to inadequately controlled population stratification. And so cultural variation and the hidden cluster problem is pernicious.
Hidden Cluster Problem
Cultural clusters (or segregated diversity) typically created by barriers impeding cultural transmission, such as topography, cultural conflict, language, social stratification by class, wealth, etc. Immigrant countries more clustered (Canada > Japan).
Countries whose borders are drawn arbitrarily with respect to the geographic arrangement of cultural groups, for example by colonial administration (many countries in Africa), are also likely to have high clustering. You can use CFST to find them: https://journals.sagepub.com/doi/abs/10.1177/0956797620916782
Note that cultural clustering is not the same as genetic clustering as we explain at length in Section 3.2.2. Indeed, reconciliation between cultural evolution and behavioral genetics requires an update in the way we think about culture.
Causal Locus Problem
Hidden cluster problem describes complexity that exists w/in social groupings. Culture is not an unstructured exogenous variable. Culture is constructive system that accumulates functional adaptations in a directed manner over time. Two key lessons here.
Lesson 1: Genes that make vs genes that break. The more complex a system, the more ways it can fail. Take the history of lighting.
Wood fire can be extinguished in 2 ways Flourescent bulbs have 7 ways to fail LEDs have 30
Faulty O-ring can explode a space shuttle and so on.
There is a fundamental asymmetry: easier to find ways to break the system than ways to explain or improve it. So too for gene function. All your cells have the same bootstrapped code, but they interact with each other, what they create, and their surroundings to create you.
There are many ways these interactions can go wrong. It is easier to identify deleterious genetic mutations than beneficial mutations. The space of failure is larger than the space of success, making genes that break more detectable than genes that make. For example, a single mutation can cause Mendelian disorders such as cystic fibrosis and Huntington’s disease, but no single mutation creates genius. Over 1000 genes have been linked to intelligence.
Each gene only explains a miniscule fraction of variation in intelligence, and the causal mechanisms are unlikely to be straightforward. In contrast to these genes that make, the causal mechanisms behind single gene mutations that cause intellectual disability—e.g. BCL11A, PHF8, ZDHHC9—are relatively well understood.
Increasing nutrition, improving schooling, and removing parasites have positive effects on IQ, but in a society where parasite infection is kept under control, we would not notice that parasite status correlates with intelligence. And by corollary, genes that provide protection against malnutrition, parasites, or pollution would only be positively associated with intelligence in environments where these insults occur. In environments where these insults have been removed, the same genes would not be associated with intelligence, and can even be deleterious, as with sickle cell trait. Not helpful if there’s no malaria.
Genes are functionally masked by cumulative cultural evolution, and we expect that this masking is extensive and systematic. A quick evolutionary and historical example: Vitamin C, the GLO gene, and dead sailors.
Vitamin C is an essential nutrient and its acquisition is thereby an essential biological function. Endogenous synthesis of vitamin C requires a gene called GLO, and GLO is present across most of the animal kingdom. But because vitamin C synthesis is metabolically costly, the gene is inactive in some species that have access to sufficient quantities of the nutrient in their diets. e.g. taxa such as teleost fishes, guinea pigs, many bats, some passerine birds, monkeys and apes.
Anthropoid primates occupy a frugivorous niche, and fruits often contain sufficient vitamin C. Here gene function is offloaded onto environmental resources. In turn, this offloading has behavioral implications. If a species becomes dependent on its environment for vitamin C, both its behavioral range and evolutionary trajectory become constrained by the availability of the nutrient. Humans are a nice example of this.
As our species migrated across the planet, we found ourselves in environments where vitamin C was in short supply. A deficiency of vitamin C causes scurvy—the bane of seafarers until the trial-and-error discovery that certain food items like sauerkraut and citrus could prevent ships from being packed with tired, bleeding, toothless, and eventually dead sailors.
Masking does not necessarily need to be in the direction from culture to genes: genetic assimilation is same process working in the opposite direction, where a trait that is regularly acquired through learning gradually transfers its locus to the genome (i.e. Baldwin effect).
Cultural Simpson’s Paradox
Which leads us to the Cultural Simpson’s Paradox. Causal Locus Problem can confound the measurement of genetic effects due to Hidden Cluster Problem obscured by WEIRD Sampling problem creating a Simpson’s paradox.
Let’s return to the UV example. The melanin-UV mismatch can be masked by the cultural diffusion of sunscreen, especially in regions with more exposure to sunlight. In other parts of the world, the issue is under-exposure to the sun causing vitamin D deficiency. Low vitamin D leads to lack of bone integrity, muscle strength, autoimmune disease, cardiovascular disease, cancer etc.
In US and France, more north you go, the the lower vitamin D levels. Makes sense, right?
But when we compare across Europe, we see the opposite pattern where people in northern countries have higher vitamin D than people in southern countries. What’s going on?
High consumption of fatty fish and cod liver oil in Northern Europe, as well as greater sun-seeking behavior in these countries compared to Mediterranean Europe. These are potent cultural adaptations.
Participants fed the traditional Norwegian fish dish mølje three times over a span of two days had 54 times the recommended daily dosage of vitamin D. The relationship between latitude and Vitamin D goes one way within a country, and the other way between the countries.
If we had been Martian anthropologists who did not know that the populated landmass known as “Europe” can in fact be broken down into sub-units called “countries”, these examples would be standard examples of a Simpson’s paradox.
In these cases, the paradox occurs when we do not know how to partition the higher-order population (Europe) into lower-order units. Fortunately, we can partition continents into countries, but in other cases, the relevant units is not as easily identifiable. Let’s move on.
We now have enough to make sense of puzzles in behavioral genetics such as (1) differences in heritability across socioeconomic levels, (2) differences in heritability across development, and (3) the Flynn effect.
SES: Heritability of IQ is higher among affluent, high socioeconomic status (SES) households than among poorer, low-SES households in some societies, but mixed in others. Why?
One explanation is ‘reciprocal causation’: genes well suited to a task can better nurture their skills in a wealthier environment than in a poorer environment and this is amplified over time. Maybe, but then why don’t we see the effect in Europe and Australia?
Here’s what we think is going on: in the US, the differences between school and home environments among high-SES households is smaller than among low-SES households. US is a land of variance. Factors such as school lotteries can dramatically affect the cultural input.
In contrast, the cultural environment is less unequal in western Europe and Australia, where, for example, high quality schools are available across SES. Where these two explanations make different predictions is for poorer countries.
Reciprocal causation would predict low heritability in poorer countries. We would predict high heritability where there is equal access to similarly poor schools and household conditions, but low heritability if inequality is high.
Incidentally we predict the opposite between human and animal environmental effects due to social transmission. It’s interesting, but not central. Check out Section 4.1.2. Let’s move onto heritability across development.
Heritability changes over the lifespan. Heritability of political orientation is similar for American identical and fraternal twins from middle childhood up to early adulthood. Right around the age at which American children leave home, this pattern is broken.
Drops for fraternal but not identical. We argue this is due to vertical vs oblique transmission and would predict a different drop off for say Italian or Croatian who leave home past 30.
Flynn effect describes the rise in IQ test scores over time. Largest in countries that have recently started modernizing, and smallest in countries that had attained modernization. No consensus to explain it, but given speed genes obviously unlikely.
We argue its caused by a rapid worldwide increase of cultural practices, technologies. Intelligence is about hardware—genes, parasites, pathogens, pollution, and nutrition affecting health and brain development, but also software—our increasingly complex cultural package.
By this account, not only is the idea of a culture-free IQ test implausible, but so too is the idea of culture-free IQ. Lots to say here. Go read Section 4.3.
Home stretch: Cultural Evolutionary Behavioral Genetics. The thrust of our theoretical case is that human psychology and behavior have a large cultural component that has been changing over history.
Most recently our psychology has been shaped by the advent of writing, numeracy, different types of agriculture, the Industrial Revolution, the Internet, and smart phones.
As new adaptive traits emerge, initially those who possess these traits will have an advantage, as in the case of access to new food sources, better healthcare, more efficient technologies, or easier methods of learning.
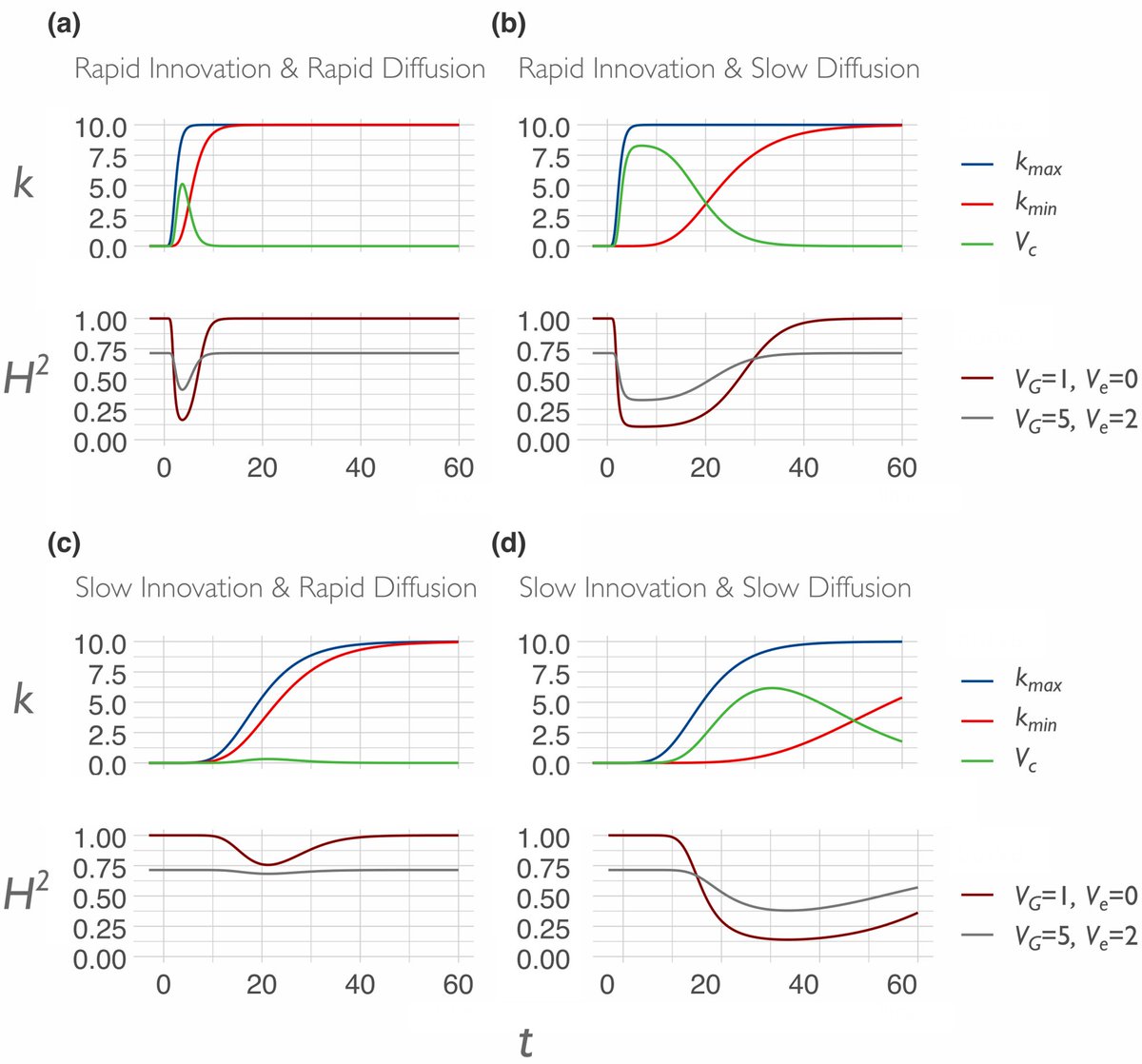
But eventually the traits will reach fixation in the population through the processes of cultural diffusion, at least until they are unseated by subsequent innovations. We predict that these cultural dynamics are reflected in heritability estimates.
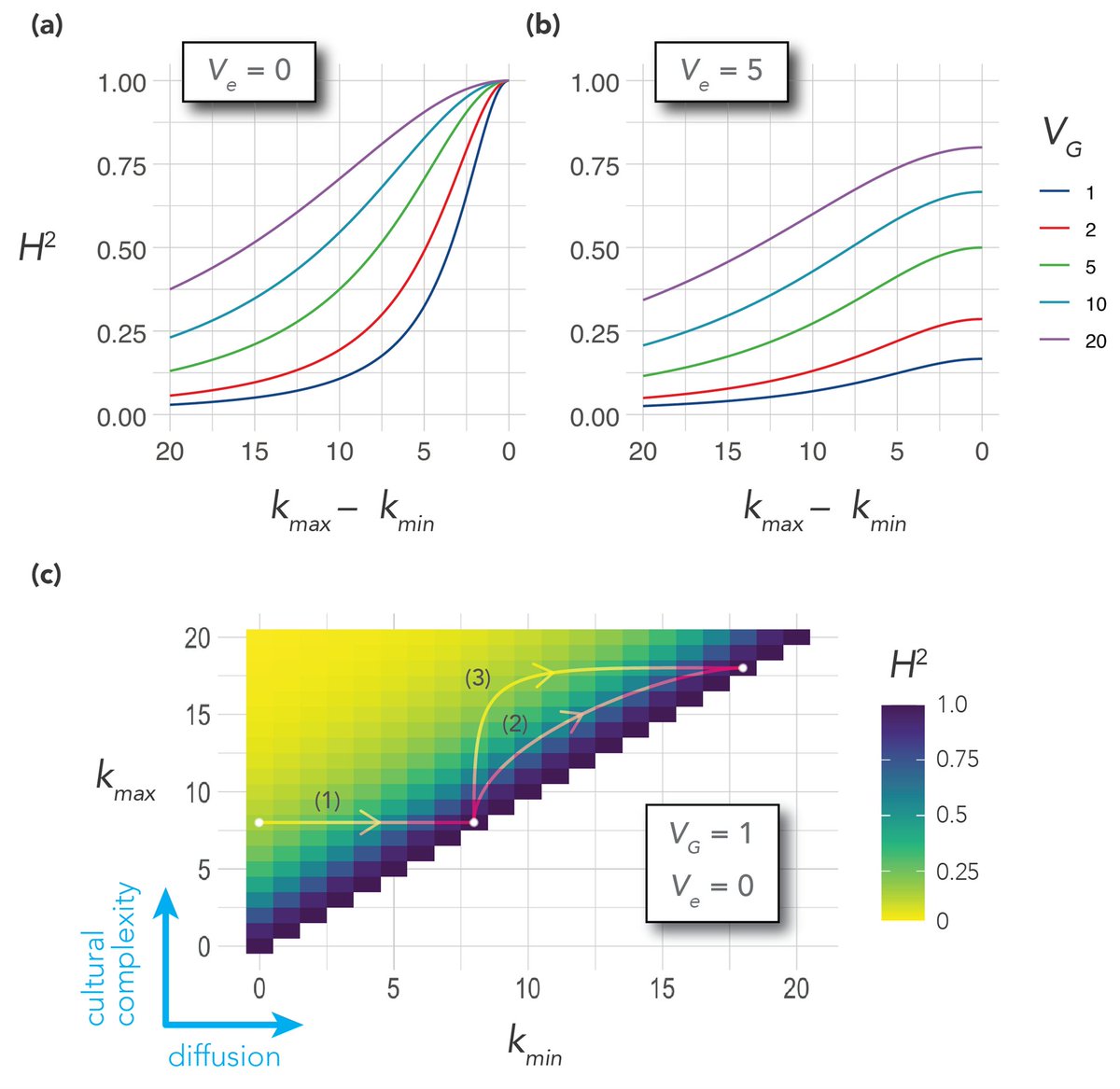
As any geneticist knows, genetic heritability is a function of the variability in the environment, variability in genes, and variability in the phenotype. There is little to predict if the phenotype is homogenous, as in the number of fingers or kidneys.
There is little to predict with if the environment or genes are homogenous. But what is factored into the environment includes not only the physical ecology, but also the cultural environment.
While variance in genes and ecology may be relatively stable, the variance in the cultural environment is continually changing through the processes of cultural evolution. A genetic account of human psychology and behavior must also account for culture and cultural evolution.
Section 5 and the conclusion tie everything together, but I’ll leave you to read it (muth.io/cegh). There’s a formal model with some pretty graphs in the Appendix:
I spent the last few days at the Templeton World Charity Foundation (TWCF) Grantee Meeting, online instead of the University of St Andrews, Scotland (due to the pandemic). This conference has become an annual highlight for me, I thoroughly enjoy discussing intelligence with an amazingly diverse and interdisciplinary group of brilliant scholars.
This year’s Diverse Intelligences Summer Institute was held online due to the pandemic. I delivered a lecture over Zoom on “What affects our level of intelligence?” followed by a lively discussion with the students. The lecture discussed brain evolution, the Cultural Brain Hypothesis, collective brain, and a cultural evolutionary account of intelligence. You can watch it below:
I spent the last few days as a meeting organized by the Templeton World Charity Foundation (TWCF) at the Diverse Intelligences Summer Institute Meeting at University of St Andrews, St Andrews, Scotland. It was an amazing few days of stimulating conversation on intelligence.
To very briefly summarize, innovation is often assumed to be an individual endeavor driven by geniuses and then passed on to the masses. Consider Thomas Edison and the lightbulb or Gutenberg and the printing press. We argue that rather than a result of far-sighted geniuses, innovations are an emergent property of our species’ cultural learning abilities, applied within our societies and social networks. Our societies and social networks act as collective brains.
Innovations, large or small, do not require heroic geniuses any more than your thoughts hinge on a particular neuron.
We argue that rates of innovation are heavily influenced by:
sociality
transmission fidelity, and
cultural variance.
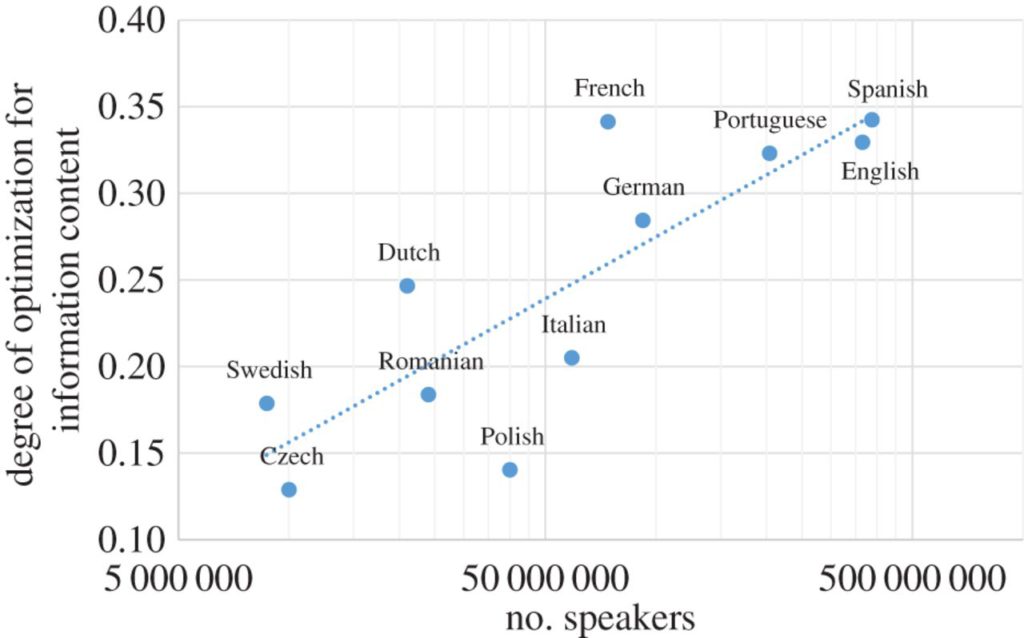
We discuss some of the forces that affect these factors. These factors can also shape each other. For example, we provide preliminary evidence that transmission efficiency is affected by sociality—languages with more speakers are more efficient.
We argue that collective brains can make each of their constituent cultural brains more innovative. This perspective sheds light on traits, such as IQ, that have been implicated in innovation. A collective brain perspective can help us understand otherwise puzzling findings in the IQ literature, including group differences, heritability differences, and the dramatic increase in IQ test scores over time.